
Review Article
*Corresponding author: Brett J Ferguson, Australian Research Council Centre of Excellence for Integrative Legume Research, The University of Queensland, St. Lucia, Brisbane, Queensland 4072, Australia, Tel: +617-3346 9951; Fax: +617-3365 3556; E-mail: b.ferguson1@uq.edu.au
Citation: van Hameren B, Hayashi S, Gresshoff PM, Ferguson BJ. Advances in the Identification of Novel Factors Required in Soybean Nodulation, a Process Critical to Sustainable Agriculture and Food Security. J Plant Biol Soil Health 2013;1(1): 6.
Copyright ©Copyright © 2013 van Hameren B, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Biology & Soil Health | ISSN: 2331-8996 | Volume: 1, Issue: 1
Submission: 07 June 2013 | Accepted: 21 June 2013 | Published: 24 June 2013
 Once the root-derived CLE peptide signal has been perceived, a novel Shoot-Derived Inhibitor (SDI) is produced which travels to the roots, presumably via the phloem, where it inhibits further nodulation [19,83]. Although SDI has yet to be identified, it has been shown to be NF dependent, heat stable, small (<1KDa) and unlikely to be a protein or RNA [84,85].
Once the root-derived CLE peptide signal has been perceived, a novel Shoot-Derived Inhibitor (SDI) is produced which travels to the roots, presumably via the phloem, where it inhibits further nodulation [19,83]. Although SDI has yet to be identified, it has been shown to be NF dependent, heat stable, small (<1KDa) and unlikely to be a protein or RNA [84,85].
Advances in the Identification of Novel Factors Required in Soybean Nodulation, a Process Critical to Sustainable Agriculture and Food Security
Bethany van Hameren, Satomi Hayashi, Peter M Gresshoff and Brett J Ferguson*
- Australian Research Council Centre of Excellence for Integrative Legume Research, The University of Queensland, Australia
*Corresponding author: Brett J Ferguson, Australian Research Council Centre of Excellence for Integrative Legume Research, The University of Queensland, St. Lucia, Brisbane, Queensland 4072, Australia, Tel: +617-3346 9951; Fax: +617-3365 3556; E-mail: b.ferguson1@uq.edu.au
Citation: van Hameren B, Hayashi S, Gresshoff PM, Ferguson BJ. Advances in the Identification of Novel Factors Required in Soybean Nodulation, a Process Critical to Sustainable Agriculture and Food Security. J Plant Biol Soil Health 2013;1(1): 6.
Copyright ©Copyright © 2013 van Hameren B, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Biology & Soil Health | ISSN: 2331-8996 | Volume: 1, Issue: 1
Submission: 07 June 2013 | Accepted: 21 June 2013 | Published: 24 June 2013
Abstract
Nodulation is a process of organogenesis that results from a symbiotic relationship between legume plants and soil-dwelling, nitrogen-fixing bacteria, called rhizobia. The rhizobia are housed in newly formed structures on the host roots, called nodules. Within nodules, the rhizobia fix atmospheric N2 into useable forms of nitrogen for the plant. This process is highly important to agriculture, as nitrogen is critical for plant growth and development and is typically the main component of fertilizers. Although fertilizers are effective, they are expensive and often pollute, making biological alternatives, such as legume nodulation, attractive for use in agriculture. Nodulation is regulated by the auto regulation of nodulation (AON) pathway, which enables the host plant to balance its needs between nitrogen acquisition and energy expenditure. Current research is elucidating the nodule development and AON signalling networks. Recent technological advances, such as RNA-sequencing, are revolutionizing the discovery of genes that are critical to nodulation. The discovery of such genes not only enhances our knowledge of the nodulation signalling network, but may help to underpin future work to isolate superior legume crops via modern breeding and engineering practices. Here, recent advances using the cutting-edge technique of RNA sequencing to identify new nodulation genes in soybean are discussed.Keywords
Legume; Nodule; Nitrogen-fixation; Rhizobia; Transcriptome; RNAseqGlobal Use of Nitrogen Fertiliser
Approximately half of the world’s population is directly reliant upon nitrogen fertiliser use in agriculture for their food supply [1-3]. Taking into account its manufacture, transport and application, the amount of fossil fuel accounts for 50% of fossil fuel use in agriculture, and 5% of the global natural gas consumption annually [4,5]. With the rising cost of fossil fuels, the use of nitrogen fertiliser is becoming increasingly expensive for farmers and is often too expensive in developing regions of the world [6]. Not only are nitrogen fertilisers expensive, they are inefficient, with 30-50% of nitrogen fertiliser typically lost to leaching. This run off can cause the eutrophication of waterways and other significant environmental problems [7]. Nitrogen contaminated drinking water can also cause methemoglobinaemia, or “Blue-baby syndrome”, a potentially fatal condition in infants [8,9].The global use of nitrogen fertiliser has been steadily increasing in most continents (Figure 1). Worryingly, this also means an increase in NOx gases, which are released when nitrogen fertiliser is broken down. These gases contribute to the formation of ground-level ozone, which causes yield reductions. Nitrous oxide (N2O) is also emitted by breakdown of nitrogen fertilisers [9,10] and is 292 times more active as a greenhouse gas than CO2 [4]. Agriculture was the main source of anthropogenic (N2O) emissions in 2005, making up 60% of the global total [11]. The majority of these emissions resulted from the application of nitrogen fertiliser [11]. By 2050 it is estimated that global nitrogen fertiliser use will increase by 50% in an attempt to boost food production and support a rising population [10]. These numbers have experts calling for agricultural reform to diminish nitrogen fertiliser use.
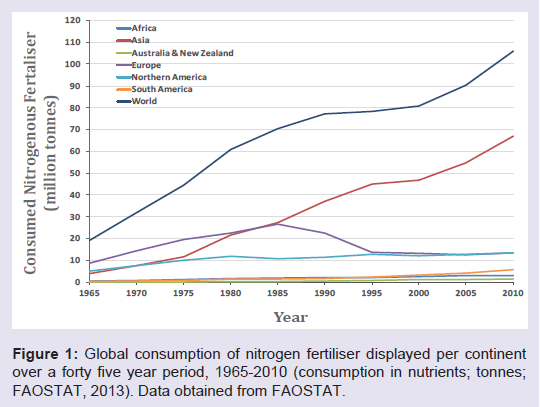
Figure 1: Global consumption of nitrogen fertiliser displayed per continentover a forty five year period, 1965-2010 (consumption in nutrients; tonnes; FAOSTAT, 2013). Data obtained from FAOSTAT.
Legume Crops as a Safe Alternative to Nitrogen Fertiliser
One safe alternative to the use of nitrogen-based fertilisers is to take advantage of biologically-fixed nitrogen. Legumes are able to form a relationship with specialized nitrogen-fixing soil bacteria, called rhizobia. The rhizobia convert atmospheric di-nitrogen into usable forms of nitrogen for the plant, whilst being housed in novel root organs, called nodules. The use of legumes as rotation crops is an important agricultural practice that many experts argue must be increased to help curb nitrogen fertiliser use [2,6,10]. Optimizing biological nitrogen fixation processes, such as nodulation, has the potential to increase crop yields and enhance soil fertility whilst simultaneously reducing farming costs and harmful environmental impacts [1,5,6,12]. However, it is only with an increase in our knowledge of nodulation processes and its genetic basis that we can fully reach this goal.Nodule Organogenesis
The most common entry point for rhizobia invasion is the region of root where the root hairs are developing, called the Zone of Nodulation (ZON) [13-16]. Rhizobia attach to the root hair, triggering root hair deformation and curling [13,14,17]. This process involves the rearrangement of underlying microtubules which allow bacterial entry and the establishment of tubular structures called the infection threads (IT) [18,19].Occurring in parallel to rhizobia invasion are inner cellular changes which lead to nodule primordia formation [19]. The ITs full of rhizobia progress towards the nodule primordia. The convergence of the rhizobia in the ITs and the nodule primordia is essential for successful nodule formation. Once the rhizobia reach the developing nodule, they are released from the ITs into specialized structures called symbiosomes, in which they differentiate into bacteroids. Using their nitrogenase enzyme complex, the rhizobia bacteroids catalyses atmospheric N2 into ammonia, which can be used by the plant [20].
Essentially, there are two different morphological types of legume nodules: indeterminate and determinate. Indeterminate nodules are initiated by inner cortical cell divisions, followed by divisions in the endodermis and pericycle, and develop persistent meristems [21-24]. Determinate nodules initially arise from cell division of the outer cortex and have transient meristems [15, 24-26].
Signalling in Nodulation
Nodule formation is initiated via a highly-specific signal exchange between compatible rhizobia bacteria and legume plants [19,27,28]. Flavonoids are released into the soil by the plant, attracting compatible rhizobia species to the host plant. They also trigger the expression of rhizobia nodulation (Nod) genes, which leads to the production of novel Nod Factor (NF) signals that are recognized by the host plant [29]. Additional rhizobia-produced factors, such as exopolysaccharides (EPS) and lipopolysaccharides (LPS), are also known to be important for nodulation and in determining rhizobiaplant specificity.Following perception of compatible partners, two main pathways are triggered within the plant: one involved in bacterial entry and infection and the other involved in cell divisions that lead to the formation of the nodule primordial [19]. Formation of a functional nodule requires synchronisation between these different signalling pathways. Many of the genes known to act in early nodulation are conserved between different legume species, but in many cases these orthologous genes have different names.
At the plant root periphery, LysM receptor kinases (Lotus japonicus Nod Factor Receptor 1 and 5; LjNFR1 and LjNFR5; soybean, Glycine max NFR1α/β and GmNFR5α/β; Medicago truncatula MtLYK3, MtLYK4 and MtNFP; and pea, Pisum sativum SYM2A and PsSYM10), perceive NF from compatible rhizobia [30-35]. These receptors are reported to associate with remorin proteins (MtSYMREM1), which may help in assembling a specialized NF receptor complex [36]. Other factors, such as LjROP6, a Rho-like small GTPase, have also been found to interact with LjNFR5 [37].
An additional receptor kinase (MsNORK/ LjSYMRK/ MtDMI2/ PsSYM19) also associates with SYMREM1 and appears to be involved in downstream signaling and possibly also the perception of NF [36]. This receptor interacts with additional nodulation factors, including a coiled-coil protein (MtRPG; [38]), a transcription factor (LjSIP1; [39]) and a 3-hydroxy-3-methylglutaryl coenzyme reductase (MtHMGR1), which is involved in isoprenoid synthesis [40]. However, the precise roles for these factors in nodulation are not yet known.
Perception of NF triggers Ca2+ fluxes, followed by the Ca2+ spiking, in root hair cells. The oscillation of Ca2+ in these cells is thought to initiate downstream signalling events [41-43]. Ca2+ spiking events require putative potassium ion-channels (MtDMI1, LjCASTOR and LjPOLLUX; [44-46]), and two nucleoporins (LjNUP133 and LjNUP85; [47,48]). Acting downstream of the Ca2+ spiking, and likely perceiving the Ca2+ signal, is a calcium and calmodulin-dependent protein kinase (CCaMK; MtDMI3/ PsSYM9; [49-52]). Novel proteins shown to interact with CCaMK include LjCIP73, which belongs to the ubiquitin superfamily [53], and MtIPD3/ LjCYCLOPS, which regulates expression of NSP1 [54-56]. Following activation of CCaMK, a number of transcription factors, including Nodulation Signalling Pathways 1 (MtNSP1) and MtNSP2 [54,57], Ets2 repressor factor (ERF), ERF required for nodulation (MtERN; [58]), and Nodule Inception (Lj/ PsNIN; [59,60]) are activated. These transcription factors work in combination to activate the expression of the early nodulation (ENOD) genes in the epidermis (e.g. MtENOD11; [61]).
CCaMK activation is also believed to trigger the increase in cytokinin level in these cells. Hormonal changes are detected by the cytokinin receptor, Lotus Histidine Kinase 1 (LjLHK1)/ MtCRE1, on the cortical cell membrane [62,63]. Activation of LjLHK1/MtCRE1 is thought to activate signalling within the cortical cells to initiate the cortical cell divisions required for the formation of the nodule [62,63].
Other components that are essential for nodule development include SCAR/WAVE proteins that appear to have roles in root hair deformation and rhizobia infection (LjNAP1/MtRIT1 and LjPIR1; [64,65]) and flotillin proteins that initiate the production of ITs for bacterial progression in the root (MtFLOT 2 and 4; [66]). An ankyrin protein that may have a role in IT development (MtVAPYRIN) is also required [67], in addition to a number of transcription factors (e.g. MtERF1 and EFD), U-box proteins (e.g. LjCERBERUS/MtLIN and MtPUB1) and early nodulin proteins of unknown function (e.g. ENOD11 and ENOD40 (reviewed in Ferguson et al. [19]). Further, a subunit of a signal peptidase complex (MtDNF1) that has a role in the processing of nodule specific cysteine-rich (NCR) proteins is an essential factor for rhizobia differentiation into nitrogen-fixing bacteroids in Medicago truncatula [68].
Autoregulation of Nodulation
The formation and maintenance of nodules is an energyintensive process. As such, the plant strictly regulates the number of nodules it forms, regardless of rhizobia availability, through the autoregulation of nodulation (AON). AON acts systemically, following nodule development. Root-derived CLV3/ESR-related (CLE) peptide hormones are synthesised, called GmRIC1 and GmRIC2 in soybean, LjCLE-RS1 in Lotus and MtCLE12/13 in Medicago [69-73]. These signals are predicted to travel to the shoot, presumably via the xylem [72,74], where they are thought to be perceived by an LRR receptor kinase, called GmNARK/LjHAR1/MtSUNN/PsSYM29 [75-78]. Mutants lacking a functional version of GmNARK/LjHAR1/MtSUNN/PsSYM29 are unable to regulate their nodule numbers and exhibit a super- or hyper-nodulating phenotype (Figure 2). It is possible that this receptor acts in conjunction with other receptor components, such as Lj/PsCLAVATA2 and/or LjKLAVIER [79,80]. Three additional factors, two Kinase-Associated Protein Phosphatases, GmKAPP1 and GmKAPP2 [81] and a putative Ubiquitin Fusion Degradation protein, GmUFD1a [82] have also been shown to possibly interact with GmNARK as part of the AON pathway.
Figure 2: GRoot systems of wild-type (WT) and supernodulating mutant (nod++) soybean plants exhibiting mature nodule structures as a result of a symbiotic relationship with Bradyrhizobium japonicum.
Nodulation is not only regulated by the number of nodulation events, but also in response to environmental factors such as stress (e.g. ethylene), soil acidity and soil nitrate (e.g. [86-92]). This gives the host plant the ability to regulate nodule development in response to its surrounding environment, thus optimizing nodulation and nitrogen-fixation under a variety of growing conditions.
The New Generation of Gene Discovery: RNA-seq
The identification of factors acting in the development and control of legume nodules has considerably increased our understanding of these processes. Moreover, it has provided novel targets for breeding and engineering programs dedicated to generating superior crop species. Recent technological advances have significantly increased the speed and efficiency with which new molecular components can be discovered. This includes new, high-throughput sequencing technology that has enabled the genomes of many legume species to be assembled in recent years, including soybean, L. japonicus, M. truncatula, chickpea and pigeon pea [93-97]. Similarly, nextgeneration RNA-sequencing (RNA-seq) technology enables the complete transcriptome of a given plant sample to be determined. This includes establishing the expression of both known and unknown genes in a sample. These benefits cannot be achieved using other techniques, such as microarrays.Soybean is one legume species that has recently been subjected to a number of RNA-seq studies seeking to identify new factors required for nodulation within its transcriptome. Indeed, soybean is often used as a model legume species [92,98], as it has had its complete genome sequenced [94], with gene atlases and gene expression databases also being publically available [99-101]. It is also amenable to a number of molecular approaches, including Agrobacterium rhizogenes-mediated transformation (e.g., [102,103]), that are essential for follow-up research aimed at confirming and functionally characterising the role of candidate genes in nodulation. In addition, a number of mutant and TILLING populations are also available, which can considerably assist genetic studies [86,104-106]. Physiologically, soybeans are also excellent for scientific purposes, being fast growing, high yielding, amenable to grafting and of appropriate size for most field and laboratory studies [6].
Soybean research is highly applicable to other legume crops including pea, lentil, chickpea, bean, peanut, lucerne, clover and faba bean [107]. Soybean is also an important crop in its own right, with production of ~250 million tonnes globally in 2011, accounting for 50% of the world’s oilseed production. It generates 200 kg N ha-1 in aboveground biomass each growing season with 58-68% of its nitrogen content resulting from symbiotic nitrogen fixation [1,2,108]. As a rotation crop, or “green manure”, soybean can be ploughed back into the soil whether their seed has been harvested or not. This provides farmers with some flexibility and helps to replenish the soil nitrogen content, as in addition to the above ground biomass, the roots and nodules contain 30-60% of the overall plant nitrogen content [109,110].
Three separate studies have reported using RNA-seq to identify differentially-expressed genes in the transcriptome of rhizobia inoculated soybean roots [16,99,111]. The genes identified represent candidates required for nodule development. An additional study used RNA-seq to determine the transcriptome of soybean leaves to identify differentially-expressed gene candidates acting in AON [82].
The work of both Libault et al. [99] and Hayashi et al. [16] focused on the early stages of nodulation, with samples harvested 48 hours post inoculation. Libault et al. [99] focused on root hairs and stripped roots, enabling a tissue-specific analysis to be conducted. In contrast, Hayashi et al. [16] focused on the ZON of the tap root, enabling nodulation-specific transcripts to be concentrated by removing transcripts found throughout the remaining portion of the root system that are not specifically nodulation related. These studies both identified a number of new nodulation gene candidates. One such candidate, GmNMNa, has already been followed up and confirmed to be involved in the regulation of rhizobia infection [112].
Barros de Carvalho et al. [111] also investigated the transcriptome of soybean root tissue; however, these authors focused their study on whole root systems harvested 10 days after rhizobia inoculation. The expression data from these samples showcase genes involved later in the nodulation pathway, including those involved in nodule maturation and growth.
To identify novel components functioning in the AON pathway, Reid et al. [82] used RNA-seq to determine the transcriptome of soybean leaves. Leaf tissue was collected from soybean shoots fed with xylem sap taken from soybean plants that were either nodulating or non-nodulating. This led to the identification of the putative ubiquitin fusion degradation protein, GmUFD1a, whose product may interact with GmNARK in the regulation of legume nodulation.
Conclusion
Collectively, the four abovementioned datasets have led to the identification of numerous new gene candidates potentially having roles in the development and regulation of soybean nodules. The confirmation and subsequent functional characterization of these genes aids in the understanding of the signalling mechanisms involved in legume nodulation. Moreover, the identification of critical nodulation genes could one day help to benefit the isolation of superior cultivars for use in agriculture and help to reduce the overapplication of nitrogen fertilisers in agriculture.Acknowledgements
This research was supported by the Australian Research Council (Centre of Excellence grant no. CEO348212 and Discovery Project grant nos. DP130103084 and DP130102266) and the University of Queensland (strategic funds and research scholarship support for BvH and SH).References
- Peoples MB, Brockwell J, Herridge DF, Rochester IJ, Alves BJR, et al. (2009) The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 48: 1-17.
- MJensen E, Peoples M, Boddey R, Gresshoff P, Hauggaard-Nielsen H, et al. (2012) Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agronomy for Sustainable Development 32: 329-364.
- Erisman JW, Sutton MA, Galloway J, Klimont Z, Winiwarter W (2008) How a century of ammonia synthesis changed the world. Nature Geoscience 1: 636-639.
- Crutzen PJ, Mosier AR, Smith KA, Winiwarter W (2007) N2O release from agro-biofuel production negates global warming reduction by replacing fossil fuels. Atmospheric Chemistry and Physics 7: 11191-11205.
- Canfield DE, Glazer AN, Falkowski PG (2010) The evolution and future ofEarth’s nitrogen cycle. Science 330: 192-196.
- Ferguson BJ (2013) The development and regulation of soybean nodules. In: A Comprehensive Survey of International Soybean Research - Genetics, Physiology, Agronomy, and Nitrogen Relationships. Ed: Board JE pp: 31-47.
- Vance CP (2001) Symbiotic nitrogen fixation and phosphorus acquisition.Plant nutrition in a world of declining renewable resources. Plant Physiol 127: 390-397.
- Murphy AP (1991) Chemical removal of nitrate from water. Nature 350: 223-225.
- Knobeloch L, Salna B, Hogan A, Postle J, Anderson H (2000) Blue babies and nitrate-contaminated well water. Environ Health Perspect 108: 675-8.
- Harvey SD, Sutton MA, Bleeker A (2013) Environmental science: The shape of nitrogen to come. Nature 494: 435-437.
- Reay DS, Davidson EA, Smith KA, Smith P, Melillo JM, et al. (2012) Global agriculture and nitrous oxide emissions. Nature Climate Change 2: 410-416.
- Hirel B, Le Gouis J, Ney B, Gallais A (2007) The challenge of improving nitrogen use efficiency in crop plants: towards a more central role for genetic variability and quantitative genetics within integrated approaches. J Exp Bot 58: 2369-87.
- Bhuvaneswari TV, Bhagwat AA, Bauer WD (1981) Transient Susceptibilityof Root Cells in Four Common Legumes to Nodulation by Rhizobia. Plant Physiol 68: 1144-1149.
- Bhuvaneswari TV, Solheim B (1985) Root hair deformation in the white clover/Rhizobium trifolii symbiosis. Physiologia Plantarum 63: 25-34.
- Calvert HE, Pence MK, Pierce M, Malik NSA, Bauer WD (1984) Anatomical analysis of the development and distribution of Rhizobium infections in soybean roots. Can J Bot 62: 2375-2384.
- Hayashi S, Reid DE, Lorenc MT, Stiller J, Edwards D, et al. (2012) Transient Nod factor-dependent gene expression in the nodulation-competent zone of soybean (Glycine max [L.] Merr.) roots. Plant Biotechnol J 10: 995-1010.
- Yao PY, Vincent JM (1969) Host specificity in the root hair “curling factor” of Rhizobium spp. Aust J Bio Sci 22: 413-424.
- Gage DJ (2004) Infection and invasion of roots by symbiotic, nitrogen-fixingrhizobia during nodulation of temperate legumes. Microbiol Mol Biol Rev 68: 280-300.
- Ferguson BJ, Indrasumunar A, Hayashi S, Lin MH, Lin YH, et al. (2010)Molecular analysis of legume nodule development and autoregulation. J Integr Plant Bio 52: 61-76.
- Udvardi MK, Day DA (1997) Metabolite transport across symbiotic membranesof legume nodules. Annu Rev Plant Physio Plant Mol Biol 48: 493-523.
- Bond L (1948) Origin and development morphology of root nodules of Pisum sativum. Bot Gaz 109: 411-434.
- Libbenga KR, Iren F, Bogers RJ, Schraag-Lamers MF (1973) The role ofhormones and gradients in the initiation of cortex proliferation and nodule formation in Pisum sativum L. Planta 114: 29-39.
- Newcomb W (1976) A correlated light and electron microscopic study of symbiotic growth and differentiation in Pisum sativum root nodules. Can J Bot 54: 2163-2186.
- Newcomb W, Sippell D, Peterson RL (1979) The early morphogenesis of Glycine max and Pisum sativum root nodules. Can J Bot 57: 2603-2616..
- Turgeon BG, Bauer WD (1982) Early events in the infection of soybean by Rhizobium japonicum. Time course and cytology of the initial infection process. Can J Bot 60: 152-161.
- Mathews A, Carroll BJ, Gresshoff PM (1989) Development of Bradyrhizobiuminfections in supernodulating and non-nodulating mutants of soybean (Glycine max [L.] Merrill). Protoplasma 150: 40-47.
- Ferguson BJ, Mathesius U (2003) Signaling interactions during nodule development. J Plant Growth Regul 22: 47-72.
- Hayashi S, Gresshoff P, Ferguson BJ (2013) Systemic Signalling In Legume Nodulation: Nodule Formation And Its Regulation. In: Baluska F. Springer-Verlag H ed. Long-distance signalling and communication in plants, signaling and communication in plants 19 ed. Germany.
- Denarie J, Debelle F, Prome JC (1996) Rhizobium lipo-chitooligosaccharide nodulation factors: signaling molecules mediating recognition and morphogenesis. Annu Rev Biochem 65: 503-35.
- Limpens E, Franken C, Smit P, Willemse J, Bisseling T, Geurts R (2003) LysM domain receptor kinases regulating rhizobial nod factor-induced infection. Science 302: 630-633.
- Madsen EB, Madsen LH, Radutoiu S, Olbryt M, Rakwalska M, et al. (2003)A receptor kinase gene of the LysM type is involved in legume perception of rhizobial signals. Nature 425: 637-40.
- Radutoiu S, Madsen LH, Madsen EB, Felle HH, Umehara Y, et al. (2003) Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 425: 585-92.
- Arrighi JF, Barre A, Ben Amor B, Bersoult A, Soriano LC, et al. (2006) TheMedicago truncatula lysin motif-receptor-like kinase gene family includes NFP and new nodule-expressed genes. Plant Physiol 142: 265-79.
- Indrasumunar A, Kereszt A, Searle I, Miyagi M, Li D, et al. (2010) Inactivationof duplicated nod factor receptor 5 (NFR5) genes in recessive loss-of-function non-nodulation mutants of allotetraploid soybean (Glycine max L. Merr.). Plant and Cell Physiol 51: 201-14.
- Broghammer A, Krusell L, Blaise M, Sauer J, Sullivan JT, et al. (2012)Legume receptors perceive the rhizobial lipochitin oligosaccharide signal molecules by direct binding. Proc Natl Acad Sci U S A 109: 13859-13864..
- Lefebvre B, Timmers T, Mbengue M, Moreau S, Herve C, et al. (2010) Aremorin protein interacts with symbiotic receptors and regulates bacterial infection. Proc Natl Acad Sci U S A 107: 2343-2348.
- Ke D, Fang Q, Chen C, Zhu H, Chen T, et al. 2012) The small GTPase ROP6 interacts with NFR5 and is involved in nodule formation in Lotus japonicus. Plant Physiol 159: 131-43.
- Arrighi J-F, Godfroy O, de Billy F, Saurat O, Jauneau A, et al. (2008) TheRPG gene of Medicago truncatula controls Rhizobium-directed polar growth during infection. Proc Natl Acad Sci U S A 105: 9817-9822.
- Zhu H, Chen T, Zhu M, Fang Q, Kang H, et al. (2008) A novel ARID DNAbinding protein interacts with SymRK and is expressed during early nodule development in Lotus japonicus. Plant Physiol 148: 337-347.
- Kevei Z, Lougnon G, Mergaert P, Horvath GV, Kereszt A, et al. (2007) 3-hydroxy-3-methylglutaryl coenzyme a reductase 1 interacts with NORK and is crucial for nodulation in Medicago truncatula. Plant Cell 19: 3974-89.
- Felle HH, Kondorosi E, Kondorosi A, Schultze M (1999) Elevation of thecytosolic free [Ca2+] is indispensable for the transduction of the Nod factor signal in alfalfa. Plant Physiol 121: 273-80.
- Wais RJ, Galera C, Oldroyd G, Catoira R, Penmetsa RV, et al. (2000) Genetic analysis of calcium spiking responses in nodulation mutants of Medicago truncatula. Proc Natl Acad Sci U S A 97: 13407-13412.
- Walker SA, Viprey V, Downie JA (2000) Dissection of nodulation signaling using pea mutants defective for calcium spiking induced by nod factors and chitin oligomers. Proc Natl Acad Sci U S A 97: 13413-8.
- Ane JM, Kiss GB, Riely BK, Penmetsa RV, Oldroyd GE, et al. (2004) Medicago truncatula DMI1 required for bacterial and fungal symbioses in legumes. Science 303: 1364-7.
- Imaizumi-Anraku H, Takeda N, Kawaguchi M, Parniske M, Hayashi M, et al. (2005) Host genes involved in activation and perception of calcium spiking. Plant Cell Physiol 46: S5.
- Riely BK, Lougnon G, Ane JM, Cook DR (2007) The symbiotic ion channel homolog DMI1 is localized in the nuclear membrane of Medicago truncatula roots. Plant J 49: 208-16.
- Kanamori N, Madsen LH, Radutoiu S, Frantescu M, Quistgaard EMH, et al. (2006) A nucleoporin is required for induction of Ca2+ spiking in legume nodule development and essential for rhizobial and fungal symbiosis. Proc Natl Acad Sci U S A 103: 359-364.
- Saito K, Yoshikawa M, Yano K, Miwa H, Uchida H et al. (2007) NUCLEOPORIN85 is required for calcium spiking, fungal and bacterial symbioses, and seed production in Lotus japonicus. Plant Cell 19: 610-624.
- Levy J, Bres C, Geurts R, Chalhoub B, Kulikova O, et al. (2004) A putative Ca2+ and calmodulin-dependent protein kinase required for bacterial andfungal symbioses. Science 303: 1361-1364.
- Mitra RM, Gleason CA, Edwards A, Hadfield J, Downie JA, et al. (2004) A Ca2+/calmodulin-dependent protein kinase required for symbiotic nodule development: Gene identification by transcript-based cloning. Proc Natl Acad Sci U S A 101: 4701-5.
- Miwa H, Sun J, Oldroyd GE, Downie JA (2006) Analysis of Nod-factorinducedcalcium signaling in root hairs of symbiotically defective mutants of Lotus japonicus. Mol Plant Microbe Interact 19: 914-23.
- Oldroyd GED, Downie JA (2004) Calcium, kinases and nodulation signalling in legumes. Nat Rev Mol Cell Biol 5: 566-576.
- Kang H, Zhu H, Chu X, Yang Z, Yuan S, et al. (2011) A novel interaction between CCaMK and a protein containing the scythe_N ubiquitin-like domain in Lotus japonicus. Plant Physiol 155: 1312-1324.
- Smit P, Raedts J, Portyanko V, Debellé F, Gough C, et al. (2005) NSP1 of the GRAS protein family is essential for rhizobial nod factor-induced transcription. Science 308: 1789-1791.
- Messinese E, Mun JH, Yeun LH, Jayaraman D, Rouge P, et al. (2007) A novelnuclear protein interacts with the symbiotic DMI3 calcium- and calmoduli independent protein kinase of Medicago truncatula. Mol Plant Microbe Interact 20: 912-921.
- Yano K, Yoshida S, Muller J, Singh S, Banba M, et al. (2008) CYCLOPS, a mediator of symbiotic intracellular accommodation. Proc Natl Acad Sci U S A 105: 20540-20545.
- Kalo P, Gleason C, Edwards A, Marsh J, Mitra RM, et al. (2005) Nodulation signaling in legumes requires NSP2, a member of the GRAS family of transcriptional regulators. Science 308: 1786-1789.
- Middleton PH, Jakab J, Penmetsa RV, Starker CG, Doll J, et al. (2007) An ERF transcription factor in Medicago truncatula that is essential for Nod factor signal transduction. Plant Cell 19: 1221-1234.
- Schauser L, Roussis A, Stiller J, Stougaard J (1999) A plant regulatorcontrolling development of symbiotic root nodules. Nature 402: 191-195.
- Borisov AY, Madsen LH, Tsyganov VE, Umehara Y, Voroshilova VA, et al. (2003) The Sym35 gene required for root nodule development in pea is an ortholog of Nin from Lotus japonicus. Plant Physiol 131: 1009-1017.
- Hirsch S, Kim J, Munoz A, Heckmann AB, Downie JA, et al. (2009) GRAS proteins form a DNA binding complex to induce gene expression during nodulation signalling in Medicago truncatula. Plant Cell 21: 545-557.
- Gonzalez-Rizzo S, Crespi M, Frugier F (2006) The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 18: 2680-2693.
- 63. Tirichine L, Sandal N, Madsen LH, Radutoiu S, Albrektsen AS, et al. (2007) A gain-of-function mutation in a cytokinin receptor triggers spontaneous root nodule organogenesis. Science 315: 104-107.
- Yokota K, Fukai E, Madsen LH, Jurkiewicz A, Rueda P, et al. (2009) Rearrangement of Actin Cytoskeleton Mediates Invasion of Lotus japonicus Roots by Mesorhizobium loti. Plant Cell 21: 267-284.
- Miyahara A, Richens J, Starker C, Morieri G, Smith L, et al. (2010) Conservation in function of a SCAR/WAVE component during infection thread and root hair growth in Medicago truncatula. Mol Plant Microbe Interact 23: 1553-1562.
- Haney CH, Long SR (2010) Plant flotillins are required for infection bynitrogen-fixing bacteria. Proc Natl Acad Sci U S A 107: 478-483.
- Murray JD, Muni RRD, Torres-Jerez I, Tang Y, Allen S, et al. (2011) Vapyrin,a gene essential for intracellular progression of arbuscular mycorrhizal symbiosis, is also essential for infection by rhizobia in the nodule symbiosis of Medicago truncatula. Plant J 65: 244-252.
- Wang D, Griffitts J, Starker C, Fedorova E, Limpens E, et al. (2010) A nodulespecific protein secretory pathway required for nitrogen-fixing symbiosis. Science 327: 1126-1129.
- Okamoto S, Ohnishi E, Sato S, Takahashi H, Nakazono M, et al. (2009) Nod factor/nitrate-induced CLE genes that drive HAR1-mediated systemic regulation of nodulation. Plant Cell Physiol 50: 67-77.
- Mortier V, Den Herder G, Whitford R, Van de Velde W, Rombauts S, et al.(2010) CLE peptides control Medicago truncatula nodulation locally and systemically. Plant Physiol 153: 222-237.
- Lim CW, Lee YW, Hwang CH (2011) Soybean nodule-enhanced CLE peptides in roots act as signals in GmNARK-mediated nodulation suppression. PlantCell Physiol 52: 1613-1627.
- Reid DE, Ferguson BJ, Gresshoff PM (2011) Inoculation- and nitrate-induced CLE peptides of soybean control NARK-dependent nodule formation. MolPlant Microb Interact 24: 606-618.
- Reid DE, Li D, Ferguson BJ, Gresshoff PM (2013) Structure-function analysis of the GmRIC1 signal peptide and CLE domain required for nodulation control in soybean. J Exp Bot 64: 1575-1585.
- Delves AC, Mathews A, Day DA, Carter AS, Carroll BJ, et al. (1986)Regulation of the soybean-rhizobium nodule symbiosis by shoot and root factors. Plant Physiol 82: 588-590.
- Krusell L, Madsen LH, Sato S, Aubert G, Genua A, et al. (2002) Shoot control of root development and nodulation is mediated by a receptor-like kinase. Nature 420: 422-426.
- Nishimura R, Hayashi M, Wu GJ, Kouchi H, Imaizumi-Anraku H, et al. (2002) HAR1 mediates systemic regulation of symbiotic organ development. Nature 420: 426-429.
- Searle IR, Men AE, Laniya TS, Buzas DM, Iturbe-Ormaetxe I, et al. (2003) Long-distance signaling in nodulation directed by a CLAVATA1-like receptor kinase. Science 299: 109-112.
- Schnabel E, Journet EP, de Carvalho-Niebel F, Duc G, Frugoli J (2005) TheMedicago truncatula SUNN gene encodes a CLV1-like leucine-rich repeat receptor kinase that regulates nodule number and root length. Plant Mol Biol 58: 809-822.
- Krusell L, Sato N, Fukuhara I, Koch BEV, Grossmann C, et al. (2011) The Clavata2 genes of pea and Lotus japonicus affect autoregulation of nodulation. Plant J 65: 861-871.
- Miyazawa H, Oka-Kira E, Sato N, Takahashi H, Wu GJ, et al. (2010) Thereceptor-like kinase KLAVIER mediates systemic regulation of nodulation and non-symbiotic shoot development in Lotus japonicus. Development 137: 4317-4325.
- Miyahara A, Hirani TA, Oakes M, Kereszt A, Kobe B, et al. (2008) Soybean nodule autoregulation receptor kinase phosphorylates two kinase-associated protein phosphatases in vitro. J Biol Chem 283: 25381-25391.
- Reid DE, Hayashi S, Lorenc M, Stiller J, Edwards D, et al. (2012) Identificationof systemic responses in soybean nodulation by xylem sap feeding and complete transcriptome sequencing reveal a novel component of the autoregulation pathway. Plant Biotechnol J 10: 680-689.
- Reid DE, Ferguson BJ, Hayashi S, Lin YH, Gresshoff PM (2011) Molecular mechanisms controlling legume autoregulation of nodulation. Ann Bot 108:789-795.
- Lin YH, Ferguson BJ, Kereszt A, Gresshoff PM (2010) Suppression of hypernodulation in soybean by a leaf-extracted, NARK- and Nod factordependent, low molecular mass fraction. New Phytol, 185: 1074-1086.
- Lin YH, Lin MH, Gresshoff PM, Ferguson BJ (2011) An efficient petiolefeedingbioassay for introducing aqueous solutions into dicotyledonousplants. Nat Protoc 6: 36-45.
- Carroll BJ, McNeil DL, Gresshoff PM (1985) Isolation and properties ofsoybean [Glycine max (L.) Merr.] mutants that nodulate in the presence of high nitrate concentrations. Proc Natl Acad Sci U S A 82: 4162-4166.
- Lorteau MA, Ferguson BJ, Guinel FC (2001) Effects of cytokinin on ethyleneproduction and nodulation in pea (Pisum sativum) cv. Sparkle. Physiol Plant 112: 421-428.
- Ferguson BJ, Ross JJ, Reid JB (2005) Nodulation phenotypes of gibberellin and brassinosteroid mutants of pea. Plant Physiol 138: 2396-2405.
- Ferguson BJ, Wiebe EMK, Emery RJN, Guinel FC (2005) Cytokinin accumulation and an altered ethylene response mediate the pleiotropic phenotype of the pea nodulation mutant R50 (sym16). Can J Bot 83: 989-1000.
- Ferguson BJ, Foo E, Ross JJ, Reid JB (2011) Relationship between gibberellin, ethylene and nodulation in Pisum sativum. New Phytol189: 829-842.
- Lin MH, Gresshoff PM, Ferguson BJ (2012) Systemic regulation of soybean nodulation by acidic growth conditions. Plant Physiol 160: 2028-2039.
- Ferguson BJ, Lin MH, Gresshoff PM (2013) Regulation of legume nodulation by acidic growth conditions. Plant Signal Behav 8: e23426.
- Cannon SB, May GD, Jackson SA (2009) Three sequenced legume genomes and many crop species: Rich opportunities for translational genomics. Plant Physiol 151: 970-977.
- Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, et al. (2010) Genome sequence of the palaeopolyploid soybean. Nature 463: 178-183.
- Young N, Debelle F, Oldroyd G, Geurts R, Cannon S, et al. (2011) The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 480: 520-524.
- Varshney RK, Chen W, Li Y, Bharti AK, Saxena RK, et al. (2011) Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat Biotechnol 30: 83-89.
- Varshney RK, Song C, Saxena RK, Azam S, Yu S, et al. (2013) Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat Biotechnol 31: 240-246..
- Ferguson BJ, Gresshoff P (2009) Soybean as a model legume. Grain Legumes 53: 7.
- Libault M, Farmer A, Brechenmacher L, Drnevich J, Langley RJ, et al. (2010)Complete transcriptome of the soybean root hair cell, a single-cell model, and its alteration in response to Bradyrhizobium japonicum infection. Plant Physiol 152: 541-552.
- Libault M, Farmer A, Joshi T, Takahashi K, Langley RJ, et al. (2010) An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J 63: 86-99.
- Severin AJ, Woody JL, Bolon YT, Joseph B, Diers BW, et al. (2010) RNASeq Atlas of Glycine max: a guide to the soybean transcriptome. BMC Plant Biol 10: 160.
- Kereszt A, Li D, Indrasumunar A, Nguyen CD, Nontachaiyapoom S, et al. (2007) Agrobacterium rhizogenes-mediated transformation of soybean tostudy root biology. Nat Protoc 2: 948-952.
- Lin MH, Gresshoff PM, Indrasumunar A, Ferguson BJ (2011) pHairyRed: A novel binary vector containing the DsRed2 reporter gene for visual selectionof transgenic hairy roots. Mol Plant 4: 537-545.
- Bolon YT, Haun WJ, Xu WW, Grant D, Stacey MG, et al. (2011) Phenotypic and genomic analyses of a fast neutron mutant population resource in soybean. Plant Physiol 156: 240-253.
- Cooper JL, Till BJ, Laport RG, Darlow MC, Kleffner JM, et al. (2008) TILLINGto detect induced mutations in soybean. BMC Plant Biol 8: 9.
- Batley J, Liu S, Mirzaei S, El-Mellouki T, Ferguson BJ, et al. (2013) TILLING in soybean reveals contrasting mutant phenotypes of LRR-receptor kinase genes GmNARK and GmCLAVATA1A. submitted.
- Rispail N, Kalo P, Kiss GB, Ellis THN, Gallardo K, et al. (2010) Model legumes contribute to faba bean breeding. Field Crops Res 115: 253-269.
- Salvagiotti F, Cassman KG, Specht JE, Walters DT, Weiss A, et al. (2008) Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res 108: 1-13.
- Mahieu S, Fustec J, Faure M-L, Corre-Hellou G, Crozat Y (2007) Comparison of two 15N labelling methods for assessing nitrogen rhizodeposition of pea. Plant Soil 295: 193-205.
- McNeill AM, Fillery IRP (2008) Field measurement of lupin belowground nitrogen accumulation and recovery in the subsequent cereal-soil system in a semi-arid Mediterranean-type climate. Plant Soil 302: 297-316.
- Barros de Carvalho GA, Batista JS, Marcelino-Guimarães FC, Nascimento LC, Hungria M (2013) Transcriptional analysis of genes involved in nodulation in soybean roots inoculated with Bradyrhizobium japonicum strain CPAC 15. BMC Genomics 14: 153-163.
- Libault M, Govindarajulu M, Berg RH, Ong YT, Puricelli K, et al. (2011) Adual-targeted soybean protein is involved in Bradyrhizobium japonicum infection of soybean root hair and cortical cells. Mol Plant Microbe Interact 24: 1051-1060.