
Research Article
Address for Correspondence: Hasan Mukhtar, Helfaer Professor of Cancer Research Director and, Vice Chair for Research, Department of Dermatology, University of Wisconsin-Madison Medical Sciences, Center, #B-25, 1300 University Avenue, Madison, WI 53706, USA, Tel: (608) 263-3297; Fax: (608) 263-5223; E-mail: hmukhtar@wisc.edu
Citation: Shabana SM, Siddiqui IA, Khan N, Adhami V, Khan QA, et al. Anti-Proliferative and Pro-Apoptotic Effects of (-)-Epigallocatechin-3- Gallate Encapsulated in Chitosan Nanoparticles on Human Prostate Carcinoma Cells. J Pharmaceu Pharmacol. 2014;2(1): 8.
Copyright © 2014 Shabana SM, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Pharmaceutics & Pharmacology | ISSN: 2327-204X |Volume: 2, Issue: 1
Submission: 28 March 2014 | Accepted: 24 April 2014 | Published: 29 April 2014
Reviewed & Approved by: Dr. Feng Li, Assistant Professor, Department of Pharmaceutical Sciences, Hampton University, USA

Next, by using colony formation assay, Chit-nanoEGCG was observed to demonstrate a remarkable dose advantage over nonencapsulated EGCG, providing a comparable inhibition of colony formation capability at 1 and 2 μM Chit-nanoEGCG when compared with 40 and 50 μM nonencapsulated EGCG in LNCaP cells (Figure 2A). Similar dose advantage was observed in PC3 cells with 4 and 6 μM dose showing effects comparable to 80 and 100 μM of native agent (Figure 2B).

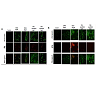
We next determined if the growth-inhibitory effects of ChitnanoEGCG correlated with enhanced induction of apoptosis. Using an Annexin-labeled fluorescent kit, we observed enhanced apoptosis of LNCaP and PC-3 cells treated with Chit-nanoEGCG for 48 h compared with high dose of native EGCG (Figure 3). We observed increased staining of annexin-V in LNCaP and PC3 cells indicating that both EGCG and Chit-nanoEGCG treatment resulted in increased apoptosis. This effect was however more prominent in Chit-nanoEGCG treated cells at a much lower doses reiterating the dose advantage of the EGCG encapsulated in nanoparticles.
To understand the molecular basis of the effects of ChitnanoEGCG, we studied several molecules involved during the initiation and execution of apoptosis. Our data showed that, levels of pro-apoptotic Bax were increased by Chit-nanoEGCG (Figure 4).
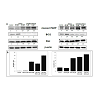
PARP (116 kDa), a nuclear enzyme that functions as a facilitator for DNA repair, is inactivated after extensive DNA damage via effector caspases during apoptosis induction; hence, inactive cleaved PARP (~85 kDa) has been profusely used a biomarker of apoptosis [25-27]. As shown in (Figure 4), we observed an increase in poly(ADP-ribose) polymerase (PARP) cleavage in both LNCaP and PC3 cells. Importantly, these responses were observed at a low concentration of nano-EGCG as compared to the native agent, further supporting a remarkable dose advantage when EGCG was delivered in nanoparticles.

Cell cycle is a series of events that controls the self-replication of mammalian cell. A normal controlled cell cycle progression is an important physiological event that is dependent on the cell efficiency to translate extracellular signaling such as mitogenic stimuli and keeps extracellular matrices undamaged in order to effectively replicate DNA and divided. This progression is regarded to be essential for normal tissue homeostasis. Loss of those controls in cells can result in unrestrained cell division which ultimately leads to cell proliferation, tumorigenesis and cancer [31]. One or more cell-cycle checkpoint defects are involved in most of the cancer types including PCa [32-34]. GTP and EGCG exhibit anti-proliferative effects versus both androgen-sensitive and androgen-insensitive human PCa cells. The effect was shown to be mediated by cell cycle deregulation and cell death induction [35]. Therefore, we next assessed the effects of Chit-nanoEGCG on cell cycle arrest in both cell lines. Induction of cell cycle (growth) arrest in S phases were reported in tumorigenic androgen independent PCa cells, while androgen dependent PCa cells were not significantly affected, even when Chit-nanoEGCG was administrated at higher doses (Figure 6) and this was associated with cyclin-dependent kinase 2 (CDK2) and cyclin E2 inhibition by chitnanoEGCG in both cell lines as compared with that seen in cells treated with EGCG and control ( Figure 7B). Chit-nanoEGCG also increased the levels of cell cycle regulatory proteins (cyclin- dependent kinase inhibitors) WAF1/p21 and CIP1/p27 proteins in both cell lines. Immunoblot analysis and the densitometric quantitation of protein bands revealed that Chit-nanoEGCG resulted in marked induction of p21 and p27 in a dose-dependent manner with a significant dose advantage over EGCG (Figure 7A).


In summary, these results demonstrate that the effectiveness of concept of nanochemoprevention (EGCG encapsulated biodegradable NPs), in terms of proapoptotic and antiproliferative efficacy, can be developed as a safe, inexpensive, tolerable and readily applicable approach for cancer control and management. The advancement in nanochemoprevention might help us to achieve higher concentrations of phytochemicals which are unattainable when the agents are provided as part of a regular diet. We suggest that, in the future this study possesses strong merit and rationale for conducting additional detailed in vivo studies in appropriate animal models with relevance to human disease.
Anti-Proliferative and Pro-Apoptotic Effects of (-)-Epigallocatechin-3- Gallate Encapsulated in Chitosan Nanoparticles on Human Prostate Carcinoma Cells
Sameh M. Shabana1,3, Imtiaz A. Siddiqui1, Naghma Khan1, Vaqar Adhami1, Qateeb A. Khan1, Shaker Mousa2, A. E. Hagras3, MA EI-Missiry3 and Hasan Mukhtar1*
1 Department of Dermatology, University of Wisconsin-Madison, 1300 University Avenue, Medical sciences center Madison, WI 53706, USA2 Pharmaceutical Research Institute at Albany, Albany College of Pharmacy, Albany, New York, USA3Department of Zoology, Faculty of Science, Mansoura University, Mansoura 35516, Egypt*Address for Correspondence: Hasan Mukhtar, Helfaer Professor of Cancer Research Director and, Vice Chair for Research, Department of Dermatology, University of Wisconsin-Madison Medical Sciences, Center, #B-25, 1300 University Avenue, Madison, WI 53706, USA, Tel: (608) 263-3297; Fax: (608) 263-5223; E-mail: hmukhtar@wisc.edu
Citation: Shabana SM, Siddiqui IA, Khan N, Adhami V, Khan QA, et al. Anti-Proliferative and Pro-Apoptotic Effects of (-)-Epigallocatechin-3- Gallate Encapsulated in Chitosan Nanoparticles on Human Prostate Carcinoma Cells. J Pharmaceu Pharmacol. 2014;2(1): 8.
Copyright © 2014 Shabana SM, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Pharmaceutics & Pharmacology | ISSN: 2327-204X |Volume: 2, Issue: 1
Submission: 28 March 2014 | Accepted: 24 April 2014 | Published: 29 April 2014
Reviewed & Approved by: Dr. Feng Li, Assistant Professor, Department of Pharmaceutical Sciences, Hampton University, USA
Abstract
(-)-epigallocatechin-3-gallate (EGCG), the major polyphenol of green tea, has shown excellent effects in preventing, as well as treating, many cancers including prostate cancer (PCa) in preclinical model systems. However its applicability to humans has met with limited success due to several issues including its limited bioavailability. We employed the use of nanotechnology to improve the outcome of cancer chemoprevention and coined the concept ‘nanochemoprevention’. Recently, we reported the synthesis, characterization and efficacy assessment of a formulation of chitosan encapsulating EGCG (Chit-nanoEGCG) for treatment of PCa in a xenograft model [1]. Here we show the efficacy of this nanoformulation in human PCa cells for inhibition of cell growth, induction of apoptosis, and inhibition of the colony formation. We further demonstrate that the encapsulated EGCG retains its mechanistic identity for induction of apoptosis. We observed that chit- nanoEGCG treatment induces apoptosis and also results in growth arrest of the cells in S phase and the cell cycle-regulatory molecules operative in the G1/S phase of the cell cycle were significantly modulated. All these effects were seen at much lower doses in chit-nanoEGCG treated cells as compared to the native agent. Collectively, chit- nanoEGCG could prove useful for preventing and/or treating human PCa.Keywords
Nanochemoprevention; PCa; EGCG; ApoptosisIntroduction
In the United States and Europe, prostate cancer (PCa) is the most common neoplasm and the second leading cause of deaths due to cancer in men [2,3]. The American Cancer Society (ACS) estimates that 233,000 new prostate cancer cases will be diagnosed in 2014, leading to nearly 30,000 deaths [4]. In order to reduce the onset and subsequent fatalities associated with PCa, there has been a marked rise in the use of natural products [5,6]. Among these natural dietary substances, (-)-epigallocatechin- 3-gallate (EGCG), a wellstudied chemopreventive agent from green tea has shown remarkable chemopreventive potential in a wide variety of cell cultures and preclinical studies [7-9].Many in vitro studies have demonstrated the cancer preventive effects of EGCG on modulation of cellular signal transduction and metabolic pathways via induction of apoptosis, suppression of cell proliferation and transformation, as well as inhibition of angiogenesis, tumor invasion and/or metastasis [7,8,10,11]. Additionally, EGCG mediated cancer preventive effects are often linked to inhibition of antioxidant activities, promotion of cell cycle arrest, induction or inhibition of certain drug metabolism enzymes, inhibition of DNA methylation, effect on miRNA expression, suppressing NFκB activation and modulation of growth factor mediated proliferation, etc. [7,9,10,12,13].
Despite these observed promising effects in preclinical studies, EGCG has only shown limited potential in clinic, and thus has met with limited success possibly due to poor systemic delivery and bioavailability [6,14]. We introduced a concept of nanochemoprevention for enhancing the outcome of EGCG for chemopreventive intervention [15,16], which involves the use of nanotechnology to improve the pharmacokinetic and pharmacodynamics profiles of natural chemopreventive agents [14,17,18]. In the first study of its kind, we encapsulated EGCG in PLGA-PEG nanoparticles and assessed its efficacy against human prostate cancer both under in vitro and in vivo conditions. In this study, we demonstrated that encapsulated EGCG, retains its biological effectiveness with an over 10-fold dose advantage in exerting its efficacy [14,19]. This idea was subsequently exploited by several laboratories worldwide and has now become an advancing field in chemoprevention research [20-24]. The last decade has witnessed rapid advancement in the development of nanochemopreventive technology with emergence of many nanoencapsulated formulations of different dietary anticancer agents [6]. In our recent study, we synthesized nanoparticles made up of natural biopolymer chitosan encapsulating EGCG (Chit-nanoEGCG), and reported that oral administration of chitosan based nanoformulated EGCG inhibited cancer cell growth in a xenograft model. Treatment with chitnanoEGCG resulted in significant inhibition of tumor growth and secreted prostate-specific antigen levels compared with EGCG alone and control groups [24].
In this study, we demonstrate that this Chit-nanoEGCG is as potent as native agent albeit at a much lower concentration. We demonstrate a significant improvement of pro-apoptotic and antiproliferative effects of EGCG encapsulated in chitosan nanoparticles.
Materials
Anti-Bax, Bcl-2, P21, P27, cyclinE2, CDK2, cleaved caspases -3,7, 8 and 9 and cleaved PARP antibodies were obtained from Cell Signaling Technology (Beverly, MA). Anti-mouse and antirabbit secondary antibody horseradish peroxidase conjugates were obtained from Cell signaling (Beverly, MA). BCA Protein assay kit was obtained from Pierce (Rockford, IL). Novex precast Tris-glycine gels were from Bio-Rad (Hercules, CA). EGCG was purchased from Sigma-Aldrich Co. (St Louis, MO). Annexin-V-FLUOS staining kit was purchased from Roche Diagnostic Corporation (Indianapolis, IN).Cell lines
Human prostate carcinoma cells LNCaP and PC3 were obtained from American Type Culture Collection (Rockville, MD). Cells were cultured in RPMI 1640 (Life Technologies, Rockville, MD) medium with 2 mmol/L L-glutamine adjusted to contain 1.5 g/L sodium bicarbonate, 4.5 g/L glucose, 10 mmol/L HEPES, and 1.0 mmol/L sodium pyruvate, and supplemented with 10% fetal bovine serum and 1% penicillin- streptomycin. The cells were maintained under standard cell culture conditions at 37°C and 5% CO2 in a humid environment.
Cell viability assay
The effect of EGCG and Chit-nanoEGCG on the viability of cells was determined by 3- (4,5-dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Briefly, 5 × 103 LNCaP and PC3 cells were plated per well in 24 well plates in 1 ml complete culture medium. LNCaP cells (60% confluent) were treated for 48 h with EGCG 30, 40, 50 μM and Chit-nanoEGCG 2, 4, 8, and 10 μM. PC3 cells (60% confluent) were treated for 48 h with EGCG 40, 80, and 100 μM and Chit-nanoEGCG 2,4,6,8 μM. After which the medium was removed and cells were washed with PBS and incubated for 2 h with MTT reagent (300 μL, 0.5 mg/ml final concentration in medium).
The MTT solution was removed and the formazan crystals were dissolved in DMSO (300 μL), plates were shaken, and absorbance was recorded at 540 nm on a microplate reader. Experiment was repeated three times with similar results. The effect of each compound on growth inhibition was assessed as percentage of cell viability in which untreated controls were considered as 100% viable. IC50 of ChitnanoEGCG and native EGCG was calculated from the cell viability data.
Colony formation assay
Cells (8 × 103) were seeded in tissue culture dishes and treated with desired EGCG and Chit-nanoEGCG formulation. Media was replenished every 3 days with the treatments. The cells were maintained under standard cell culture conditions at 37°C and 5% CO2 in a humid environment. Colonies that formed in 2 to 3 wk were fixed with 10% buffered formalin, stained with 2% gentian violet (w/v methanol solution), washed with water, and air-dried.
Apoptosis detection by fluorescence microscopy
The Annexin V-FLUOS Staining Kit (Roche Applied Biosciences, Mannheim, Germany) was used for the detection of apoptotic cells according to the manufacturer’s protocol. The cells, LNCaP and PC3 were grown to 50-60% confluency on cell culture slides and then treated with EGCG (40, 50 μM) and Chit-nanoEGCG (1, 2 μM) and EGCG (80, 100 μM) and Chit-nanoEGCG (4, 6 μM) for 48 h. Cells that were used as controls were incubated with the culture medium only. The cells were then incubated with Annexin V- FLUOS labeling reagent for 10 min and then analyzed using fluorescence microscopy (excitation at 450-500 nm and detection at 515-565 nm). Cells with green fluorescence were scored as apoptotic and the fluorescence was visualized using a Nikon Eclipse Ti system (Nikon Instruments Inc., Tokyo, Japan). Digital images were captured with an attached Cool 1SNAP camera (Roper Scientific, Trenton, NJ) linked to a computer.
Protein extraction and Western blotting
The cells were treated with EGCG and Chit-nanoEGCG in complete medium. 48 hours later, the cells were washed with cold PBS (10 mmol/L, pH 7.4) followed by incubation in ice-cold lysis buffer [50 mM Tris-HCl, 150 mM NaCl, 1 mM ethyleneglycol-bis(aminoethylether)-tetraacetic acid, 1 mM ethylenediaminetetraacetic acid, 20 mM NaF, 100 mM Na3VO4, 0.5% NP-40, 1% Triton X-100, 1 mM phenylmethylsulfonyl fluoride, pH 7.4] with freshly added protease inhibitor cocktail (Protease Inhibitor Cocktail Set III, Calbiochem, La Jolla, CA) and 0.2 mM sodium vanadate over ice for 30 min. The cells were scraped and the suspension was collected in a microfuge tube and passed through a 21.5-gauge needle to break up the cell aggregates. The lysate was cleared by centrifugation at 14,000 × g for 15 min at 4°C, and the supernatant (total cell lysate) was stored at -80°C for further analysis. The protein concentration was determined by BCA protein assay kit according to the manufacturer’s protocol.
For immunoblotting, 30-50 μg protein was resolved over 8-12% polyacrylamide gels and transferred to a nitrocellulose membrane. The blot was blocked in blocking buffer (5% non-fat dry milk/1% Tween 20 in 20mM Tris-buffered saline, pH 7.6) for 1h at room temperature, incubated with appropriate monoclonal or polyclonal primary antibody in blocking buffer for 1.5 h to overnight at 4°C, followed by incubation with anti-mouse or anti-rabbit secondary antibody horseradish peroxidase (HRP) conjugate obtained from Cell Signaling technologies. The membrane was washed and bound complex detected by chemiluminescence (ECL kit; Amersham Biosciences, UK) and autoradiography using Bio-Rad Gel-Doc (Bio-Rad Laboratories). Densitometric measurements of the bands in immunoblotting were performed using digitalized scientific software program Image J (National Institutes of Health).
Cell-cycle analysis by Flow cytometry
Cells treated with/without EGCG and Chit-nanoEGCG for 48 hours were processed using APO-DIRECT assay kit (BD Biosciences Pharmingen, San Diego, CA, USA). Briefly, post-treatment cells were processed for labeling with fluorescein-tagged dUTP nucleotide and propidium iodide and analyzed using Modfit software. The treated cells were fixed in 1% paraformaldehyde (methanol free) in PBS for 60 minutes on ice, centrifuged (5 min; 300 xg), washed with PBS and pelleted again. Cells were resuspended in 70% ice cold ethanol and kept on ice for 30 minutes. The cells were pelleted (5 min; 300 xg) and washed twice in the wash buffer provided in the kit. DNA labeling solution containing TdT enzyme and FITC dUTP was used to label the cells for 60 min at 37°C. Cells were again washed twice by centrifuging in wash buffer and finally the pellet was resuspended in PI/RNase staining buffer provided in the kit and analyzed using a FACSCalibur.
Statistical analysis
Results were expressed as the means ± SD. Statistical analysis between controls and treatments were performed by Student’s t-test, where a P ≤ 0.05 was considered statistically significant.
Results and Discussion
In our first set of experiments, we evaluated the effect of ChitnanoEGCG versus native EGCG on cell viability of androgensensitive LNCaP and androgen-independent PC3 human prostate cancer (PCa) cells. This choice was based on the premise that PCa at the time of diagnosis represents a mixture of androgen -dependent and -independent cells, and a treatment protocol aimed at eliminating both types of cells would be ideal. As shown in Figure 1, treatment of LNCaP and PC3 cells with EGCG and Chit-nanoEGCG for 48 h resulted in a dose-dependent inhibition of cell growth as analyzed by MTT assay. The effective dose of EGCG for inhibiting cell growth was reduced significantly when encapsulated in chitosan nanoparticles. The IC50 (at 48 h post treatment) of Chit- nanoEGCG was observed to be 4 μM as compared to 90 μM with native EGCG in PC3 cells. For LNCaP cells the IC50 of Chit-nanoEGCG was observed to be 1 μM as compared to 45 μM of the native agent. The differential response in the two cells is possibly due to the fact that the two cells (differing in androgen status) behave totally differently in presence of an agent. We speculate that nano-EGCG enters into the cells via endocytosis. They adsorb serum proteins nonspecifically onto their surface and enter the cell via receptors on the cell membrane [14].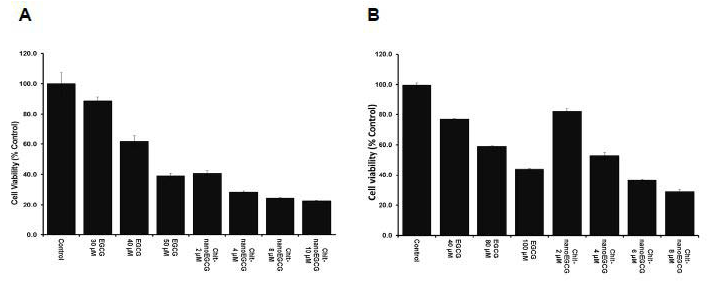
Figure 1: Comparative effects of nonencapsulated EGCG and Chit-nanoEGCG treatment on cell viability. LNCaP cells (A) and PC3 cells (B) were treated with EGCG and Chit-nanoEGCG at indicated doses for 48 h, and cell growth was determined by the MTT assay as described under Materials and Methods.
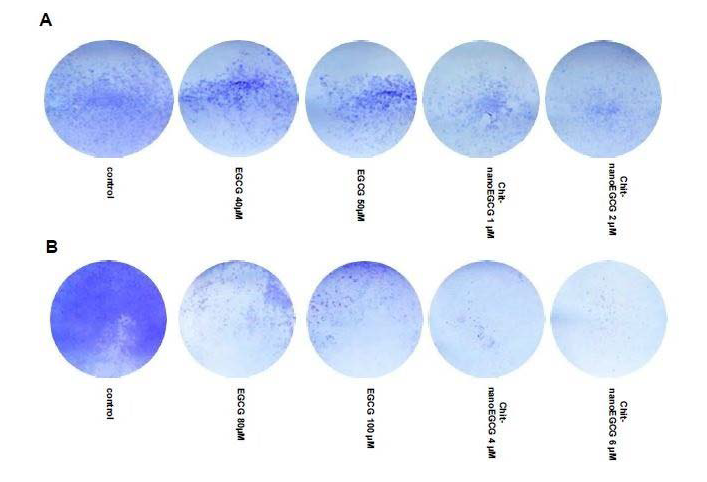
Figure 2: Comparative effects of nonencapsulated EGCG and Chit-nanoEGCG treatment on colony formation. LNCaP cells (A) and PC3 cells (B) were treated with EGCG and Chit-nanoEGCG at indicated doses for 48 h and the plates were observed for colonies. At the end of experiments colonies of the prostate cells were washed with PBS and stained with crystal violet, counted and pictured.
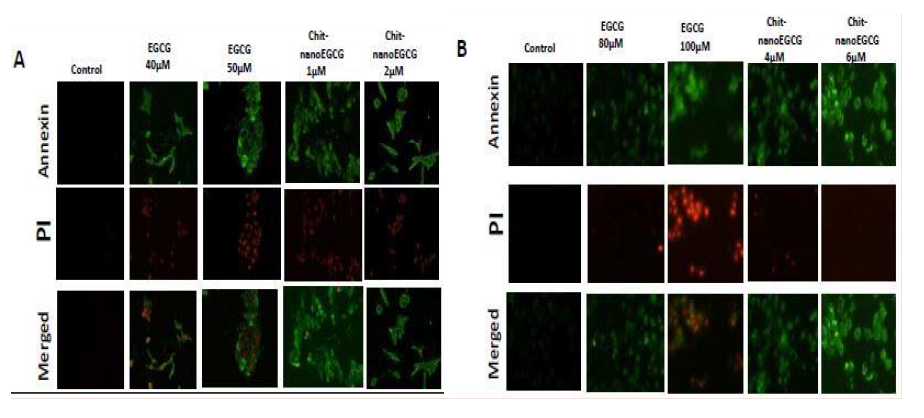
Figure 3: Comparative effects of nonencapsulated EGCG and Chit-nanoEGCG treatment on apoptosis. LNCaP cells (A) and PC3 cells (B) were grown on cell culture slides and treated with EGCG and Chit-nanoEGCG at indicated doses for 48 h. Cells with green fluorescence were scored as apoptotic. Images (magnification, 20X) shows representative pictures from two independent samples.
Concomitantly, the levels of anti-apoptotic Bcl-2 were decreased by Chit-nanoEGCG. Furthermore, modulations in the expression of key regulators of apoptosis Bax and Bcl- 2 by these agents resulted in shift in the ratio of these molecules in a way that favored apoptosis (Figure 4). These effects were observed in both tested prostate carcinoma cells. The data also suggested that the dose advantage seen in the cell viability and apoptosis assays is persistent at the protein expression level.
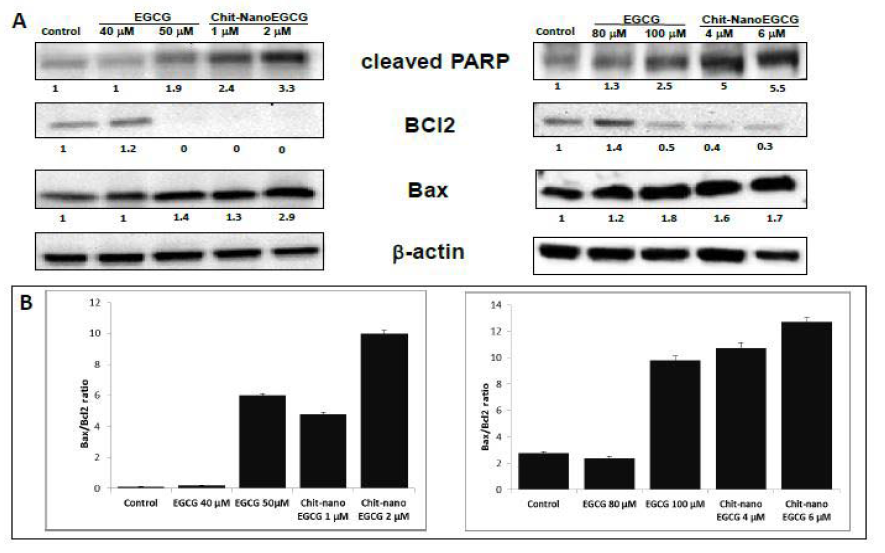
Figure 4: Comparative effects of nonencapsulated EGCG and Chit-nanoEGCG treatment on apoptotic markers: A) protein expression of Cleaved PARP, BAX and Bcl2 in LNCaP cells (left panel) and PC3 cells (right panel). B) BAX/Bcl2 ratio of LNCaP cells (left panel) and PC3 cells (right panel). As detailed in Materials and Methods, total tissue lysates were prepared and 40 μg protein was subjected to electrophoresis followed by immunoblot analysis and chemiluminescence detection. Equal loading of protein was confirmed by stripping the immunoblot and reprobing it for β-actin. The values below the figures represent relative density of the bands normalized to β-actin.
Changes in the Bax to Bcl-2 ratio always lead to the destabilization of the mitochondrial membrane and release of apoptotic factors. These factors induce the caspase proteolytic cascade and cleavage of PARP, ultimately leading to cell death. At least two pathways, the death receptor and the mitochondrial pathways, are known to activate the effector caspases via the initiator caspases [28]. The process of apoptosis consists of the sequential activation of caspases, which are the key players in apoptotic cell death [24]. Caspases 8 and 9 are initiator caspases, performing signaling functions along the apoptosis cascade, whereas caspase 3 and 7 are two of the executioner caspases that directly activate the apoptosis effectors [26,29,30]. As shown in Figure 5, we observed that there was significant increase in activation of cleaved caspases 3,7,8,9 in both tested cell lines. These proteins were over two fold higher up-regulation in the cells treated with Chit-nanoEGCG as compared to native EGCG reconfirming the dose advantage with nanoformulated EGCG. Overall, our observations suggest that the mitochondrial pathway triggers the activation of initiator caspase-8, which in turn activates the effector caspases, resulting in proteolytic cleavage of PARP.
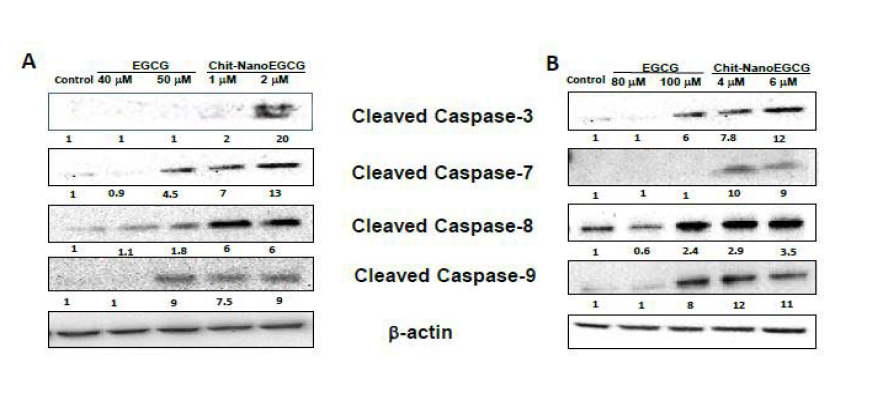
Figure 5: Comparative effects of nonencapsulated EGCG and Chit-nanoEGCG treatment on cleaved caspases. Protein expression of Cleaved Caspase-3,7,8 and 9 in LNCaP cells (A) and PC3 cells (B). As detailed in Materials and methods, total tissue lysates were prepared and 40 μg protein was subjected to electrophoresis followed by immunoblot analysis and chemiluminescence detection. Equal loading of protein was confirmed by stripping the immunoblot and reprobing it for β-actin. The values below the figures represent relative density of the bands normalized to β-actin.
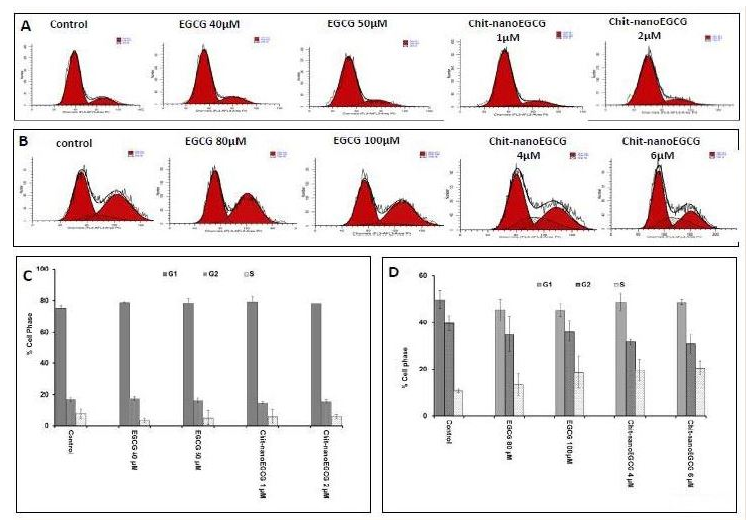
Figure 6: Flow-cytometric analysis of cell cycle. LNCaP cells (A) and PC3 cells (B) were treated with EGCG and Chit-nanoEGCG for 48 hours. Cells were sorted by FACS (fluorescence-activated cell sorting analysis), histograms of cellular DNA in G1, G2 and S phase were analyzed by ModiFitLT V3.0. Data are representative of duplicate experiments. Percentage of cell phase (G1, G2 and S) of LNCaP cells (C) and PC3 cells (D) treated with EGCG and Chit-nanoEGCG.
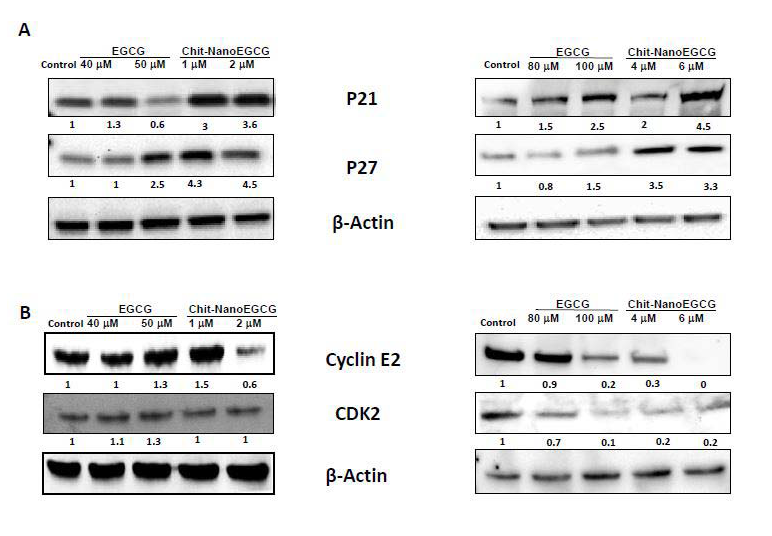
Figure 7: Comparative effects of nonencapsulated EGCG and Chit-nanoEGCG treatment on cell cycle regulatory proteins: A) protein expression of P21 and P27 in LNCaP cells (left panel) and PC3 cells (right panel); B) protein expression of cyclin E2 and CDK2 in LNCaP cells (left panel) and PC3 cells (right panel). As detailed in Materials and methods, total tissue lysates were prepared and 40 μg protein was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis followed by immunoblot analysis and chemiluminescence detection. Equal loading of protein was confirmed by stripping the immunoblot and reprobing it for β-actin. The values above the figures represent relative density of the bands normalized to β-actin.
References
- Khan N, Bharali DJ, Adhami VM, Siddiqui IA, Cui H, et al. (2014) Oral administration of naturally occurring chitosan-based nanoformulated green tea polyphenol EGCG effectively inhibits prostate cancer cell growth in a xenograft model. Carcinogenesis 35: 415-423.
- Ferlay J, Steliarova-Foucher E, Lortet-Tieulent J, Rosso S, Coebergh JW, et al. (2013) Cancer incidence and mortality patterns in Europe: estimates for 40 countries in 2012. Eur J Cancer 49: 1374-1403.
- Siegel R, Naishadham D, Jemal A (2013) Cancer statistics, 2013. CA: a cancer journal for clinicians 63: 11-30.
- Siegel R, Ma J, Zou Z, Jemal A (2014) Cancer statistics, 2014. CA Cancer J Clin 64: 9-29.
- Aggarwal BB, Takada Y, Oommen OV (2004) From chemoprevention to chemotherapy: common targets and common goals. Expert Opin Investig Drugs 13: 1327-1338.
- Muqbil I, Masood A, Sarkar FH, Mohammad RM, Azmi AS (2011) Progress in nanotechnology based approaches to enhance the potential of chemopreventive agents. Cancers 3: 428-445.
- Khan N, Mukhtar H (2013) Modulation of signaling pathways in prostate cancer by green tea polyphenols. Biochem Pharmacol 85: 667-672.
- Khan N, Adhami VM, Mukhtar H (2009) Review: green tea polyphenols in chemoprevention of prostate cancer: preclinical and clinical studies. Nutr Cancer 61: 836-841.
- Siddiqui IA, Afaq F, Adhami VM, Ahmad N, Mukhtar H (2004) Antioxidants of the beverage tea in promotion of human health. Antioxid Redox Signal 6: 571-582.
- Adhami VM, Ahmad N, Mukhtar H (2003) Molecular targets for green tea in prostate cancer prevention. J Nutr 133: 2417S-2424S.
- Siddiqui IA, Saleem M, Adhami VM, Asim M, Mukhtar H (2007) Tea beverage in chemoprevention and chemotherapy of prostate cancer. Acta Pharmacol Sin 28: 1392-1408.
- Nandakumar V, Vaid M, Katiyar SK (2011) (-)-Epigallocatechin-3-gallate reactivates silenced tumor suppressor genes, Cip1/p21 and p16INK4a, by reducing DNA methylation and increasing histones acetylation in human skin cancer cells. Carcinogenesis 32: 537-544.
- Fang MZ, Wang Y, Ai N, Hou Z, Sun Y, et al. (2003) Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res 63: 7563-7570.
- Siddiqui IA, Adhami VM, Bharali DJ, Hafeez BB, Asim M, et al. (2009) Introducing nanochemoprevention as a novel approach for cancer control: proof of principle with green tea polyphenol epigallocatechin-3-gallate. Cancer Res 69: 1712-1716.
- Siddiqui IA, Mukhtar H (2010) Nanochemoprevention by bioactive food components: a perspective. Pharm Res 27:1054-1060.
- Siddiqui IA, Adhami VM, Ahmad N, Mukhtar H (2010) Nanochemoprevention: sustained release of bioactive food components for cancer prevention. Nutr Cancer 62: 883-890.
- Shahani K, Swaminathan SK, Freeman D, Blum A, Ma L, et al. (2010) Injectable sustained release microparticles of curcumin: a new concept for cancer chemoprevention. Cancer Res 70: 4443-4452.
- Anand P, Nair HB, Sung B, Kunnumakkara AB, Yadav VR, et al. (2010) Design of curcumin-loaded PLGA nanoparticles formulation with enhanced cellular uptake, and increased bioactivity in vitro and superior bioavailability in vivo. Biochem Pharmacol 79: 330-338.
- Siddiqui IA, Adhami VM, Chamcheu JC, Mukhtar H (2012) Impact of nanotechnology in cancer: emphasis on nanochemoprevention. Int J Nanomedicine 7: 591-605.
- Bharali DJ, Siddiqui IA, Adhami VM, Chamcheu JC, Aldahmash AM, et al. (2011) Nanoparticle delivery of natural products in the prevention and treatment of cancers: current status and future prospects. Cancers (Basel) 3: 4024-4045.
- Barras A, Mezzetti A, Richard A, Lazzaroni S, Roux S, et al. (2009) Formulation and characterization of polyphenol-loaded lipid nanocapsules. Int J Pharm 379: 270-277.
- Danhier F, Ansorena E, Silva JM, Coco R, Le Breton A, et al. (2012) PLGA-based nanoparticles: an overview of biomedical applications. J Control Release 161: 505-522.
- Dube A, Nicolazzo JA, Larson I (2010) Chitosan nanoparticles enhance the intestinal absorption of the green tea catechins (+)-catechin and (-)-epigallocatechin gallate. Eur J Pharm Sci 41: 219-225.
- Khan N, Bharali DJ, Adhami VM, Siddiqui IA, Cui H, et al. (2014) Oral administration of naturally occurring chitosan-based nanoformulated green tea polyphenol EGCG effectively inhibits prostate cancer cell growth in a xenograft model. Carcinogenesis 35: 415-423.
- Malik A, Afaq S, Shahid M, Akhtar K, Assiri A (2011) Influence of ellagic acid on prostate cancer cell proliferation: a caspase-dependent pathway. Asian Pac J Trop Med 4: 550-555.
- Degterev A, Boyce M, Yuan J (2003) A decade of caspases. Oncogene 22: 8543- 8567.
- Ame JC, Spenlehauer C, de Murcia G (2004) The PARP superfamily. Bioessays 26: 882-893.
- Bozec A, Ruffion A, Decaussin M, Andre J, Devonec M, et al. (2005) Activation of caspases-3, -6, and -9 during finasteride treatment of benign prostatic hyperplasia. J Clin Endocrinol Metab 90: 17-25.
- Fuentes-Prior P, Salvesen GS (2004) The protein structures that shape caspase activity, specificity, activation and inhibition. Biochem J 384: 201-232.
- Galluzzi L, Vitale I, Abrams JM, Alnemri ES, Baehrecke EH, et al. (2012) Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ 19: 107-120.
- Evan GI, Vousden KH (2001) Proliferation, cell cycle and apoptosis in cancer. Nature 411: 342-348.
- Mercer WE (1998) Checking on the cell cycle. J Cell Biochem Suppl 30-31: 50-54.
- Fernandez PL, Hernandez L, Farre X, Campo E, Cardesa A (2002) Alterations of cell cycle-regulatory genes in prostate cancer. Pathobiology 70: 1-10.
- Davis JN, Day ML (2002) The convergence of hormone regulation and cell cycle in prostate physiology and prostate tumorigenesis. Mol Biotechnol 22: 129-138.
- Gupta S, Ahmad N, Nieminen AL, Mukhtar H (2000) Growth inhibition, cellcycle dysregulation, and induction of apoptosis by green tea constituent (-)-epigallocatechin-3-gallate in androgen-sensitive and androgen-insensitive human prostate carcinoma cells. Toxicol Appl Pharmacol 164: 82-90.