
Research Article
*Address for Correspondence: Peter F. Surai, Sumy National Agrarian University, Sumy, Ukraine E-mail: psurai@feedfood.co.uk
Citation: Surai PF, Fisinin VI. Antioxidant-Prooxidant Balance in the Intestine: Applications in Chick Placement and Pig Weaning. J Veter Sci Med. 2015;3(1): 16.
Copyright © Surai et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.Journal of Veterinary Science & Medicine | ISSN: 2325-4645 | Volume: 2, Issue: 2
Submission: 02 February 2015 | Accepted: 09 March 2015 | Published: 13 March 2015
Reviewed & Approved by: Dr. Ioannis Hadjigeorgiou, Assistant Professor, Department of Nutrition Physiology and Feeding, Agricultural University of Athens, Greece
Peroxidized PUFAs
Antioxidant-Prooxidant Balance in the Intestine: Applications in Chick Placement and Pig Weaning
Peter F. Surai1-3* and Vladimir I. Fisinin4
- 1Trakia University, Stara Zagora, Bulgaria
- 2Szent Istvan University, Godollo, Hungary
- 3Sumy National Agrarian University, Sumy, Ukraine
- 4All Russian Institute of Poultry Husbandry, Sergiev Posad, Russia
*Address for Correspondence: Peter F. Surai, Sumy National Agrarian University, Sumy, Ukraine E-mail: psurai@feedfood.co.uk
Citation: Surai PF, Fisinin VI. Antioxidant-Prooxidant Balance in the Intestine: Applications in Chick Placement and Pig Weaning. J Veter Sci Med. 2015;3(1): 16.
Copyright © Surai et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.Journal of Veterinary Science & Medicine | ISSN: 2325-4645 | Volume: 2, Issue: 2
Submission: 02 February 2015 | Accepted: 09 March 2015 | Published: 13 March 2015
Reviewed & Approved by: Dr. Ioannis Hadjigeorgiou, Assistant Professor, Department of Nutrition Physiology and Feeding, Agricultural University of Athens, Greece
Abstract
A delicate balance between antioxidants and prooxidants in cells is an important determinant of various physiological processes and maintenance of this balance is the main aim of so called an integrated antioxidant system built in the animal body. This system was developed during evolution to provide an antioxidant defence and give a chance for animals to survive in oxygenated atmosphere. Recent data suggest that the antioxidant-prooxidant balance starts in the intestine. Indeed, redox status of the gut is involved in regulations of many important physiological processes. When food is consumed and appears in the stomach and then in the small intestine, it contains a range of antioxidants (vitamins E, C, carotenoids, flavonoids, etc.) but may also contain a range of potentially dangerous substances (oxidized lipids, mycotoxins, traces of heavy metals, etc.). To keep a balance between antioxidants and prooxidants in the intestine is a very important task in which diet plays a key role. Recently a vitagene concept was developed indicating that vitagenes are responsible for synthesis of a variety of antioxidant molecules increasing adaptive ability of human/animals to various stress conditions. Furthermore, there is a range of nutrients able to affect vitagenes. In poultry production, chicken placement is considered to be a substantial stress and dietary antioxidants could have a protective effect. Similarly, in pig production, weaning process is also a great stress for piglets. Preliminary data indicates that usage of antioxidant compositions upplying with water for newly hatched chicks and newly-weaned piglets could become an important part of the animal production technologies.Introduction
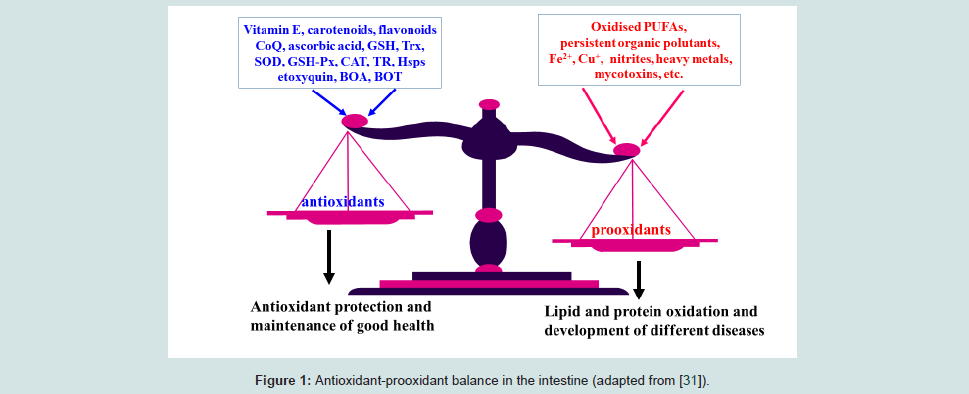
A delicate balance between antioxidants and prooxidants in cells is an important determinant of various physiological processes and maintenance of this balance is the main aim of so called an integrated antioxidant system built in the animal body. This system was developed during evolution to provide an antioxidant defence and give a chance for animals to survive in oxygenated atmosphere. Recent data suggest that the antioxidant-prooxidant balance starts in the intestine (Figure 1). Indeed, redox status of the gut is involved in regulations of many important physiological processes.The aim of this review is to update recent knowledge on antioxidant-prooxidant balance in the animal/poultry gut and consider its application in newly hatched chick and weaned piglet nutrition.
The Gastrointestinal Tract (GIT) as a Major Site of Antioxidant Action
It is well appreciated that some antioxidants, such as antioxidant enzymes including glutathione peroxidase (GSH-Px), superoxide dismutase (SOD) and catalase, glutathione (GSH), thioredoxin (Trx), coenzyme Q, can be synthesised in the body. However, the diet is the major provider of nutrients possessing antioxidant properties directly (vitamin E, vitamin A, carotenoids, ascorbic acid, flavonoids etc.) or essentials for the synthesis of antioxidant enzymes. For example, selenium (Se) is an essential part of a range of selenoproteins performing antioxidant functions (GSH-Px, thioredoxin reductase (TR), selenoproteins P and W, etc.); mangan (Mn) is an integral part of mitochondrial Mn-SOD; zinc (Zn) and copper (Cu) are integral arts of cytosolic Cu, Zn-SOD; iron (Fe) is an integral part of catalase.When food is consumed and appears in the stomach and then in the small intestine, it contains a range of antioxidants but may also contain a range of potentially dangerous substances. To keep a balance between antioxidants and prooxidants in the intestine is a very important task in which diet plays a crucial role. In general, prooxidants which can be found in the GIT could be summarised as follows:
Prooxidants in the GIT
Lipid hydroperoxides (LOOH) derived from unsaturated fatty acids are important intermediates of peroxidative reactions induced by reactive oxygen species (ROS). In fact, lipid hydroperoxides are not stable and in the presence of transition metal ions can decompose producing new free radicals and cytotoxic aldehydes [1]. Oxidised lipids are partly absorbed in the digestive tract [2] and incorporated into membrane phospholipids altering their structure and properties [3,4]. In animal models it has been shown that oxidised lipids in the diet can suppress growth [5,6], reduce vitamin E level in tissues increasing their susceptibility to lipid peroxidation [7], increase tissue protein oxidation [4] and increase the number of aberrant crypts in the intestine [8]. The consumption of oxidized fats is associated with diarrhoea, liver enlargement, growth depression and histological changes in tissues of experimental animals [9,12]. Indeed, feeding weaning pigs diets containing 10% thermally oxidized lipids for 38 d appeared to impair oxidative status [13] and negatively affected growth erformance and liver triglyceride concentration of young pigs, which was associated with an upregulation of fatty acid catabolism pathways [14]. Furthermore, the oxidized fat in the pig diet up-regulated sterol regulatory element-binding protein and its target genes in liver and small intestine [15]. The gastrointestinal epithelium of swine and chickens responded to oxidant stress imposed by oxidized fat by increased enterocyte turnover and the gut associated immune system was compromised [16]. From data presented above it is clear that lipid peroxidation in the feed is an important source of toxic products and a potential source of free radicals in the animal digestive tract.
Iron ions
Iron is recognised as an essential nutrient; however, iron absorption from a diverse diet has been shown to be about 15% [17]. The main problem with iron nutrition is its reactivity and possible involvement in free radical generation. Iron ions are considered to catalyse the formation of the hydroxyl radical and accelerate the decomposition of lipid hydroperoxides [18] and to stimulate lipid peroxidation [19,20]. Iron dietary supplementation could represent an important source of potentially dangerous free iron in the digestive tract which could also be involved in lipid peroxidation in the intestine [21]. For example, iron supplementation in relatively high doses resulted in abnormal iron accumulation and increased lipid peroxidation in rats [22]. Iron supplementation amplified the inflammatory response and enhanced the subsequent mucosal damage in a rat model of colitis [23]. In the conditions of dietary iron overload in rats, in vivo hydroxyl radical generation (ultimately responsible for iron-induced injury) was demonstrated [24]. Iron can also interact with other nutrients stimulating free radical production.For example, it has been shown that iron in combination with the secondary bile acids, lithocholic and deoxycholic acids and the vitamin K group can generate free radicals [25]. Clearly, feed iron supplementation represents a possible source of catalytic iron in the GIT responsible for oxidative stress and lipid and protein oxidation.
Nitrites and nitrates
Nitrite is consumed in the diet, through feed and drinking water [26]. Therefore, animals are subjected to significant nitrate and nitrite levels in feed and water, as well as those formed in vivo. Nitrites and nitrates formed from nitrogenous sources by microorganisms in saliva and intestine are considered to be the major source of animal and human exposure under physiological conditions [27]. It is generally accepted that nitrate is concentrated in the saliva and rapidly conversed to nitrite by facultative anaerobic bacteria. Benjamin et al. showed that nitrite is converted to NO under the highly acidic conditions (pH 3) which occur in the lumen of the stomach [28]. It was observed that the generation and accumulation of NO from typical nitrite concentrations found in biological tissues increases 100-fold when the pH falls from 7.4 to 5.5 [29]. Therefore nitrate and itrite can generate NO* radical by either direct disproportionation or reduction under the acidic and highly reduced conditions [27]. More importantly, NO* can be a source of another reactive free radical peroxynitrite (ONOO-), which is 1000 times more oxidizing than H2O2 and has a half-life in solution of about 1–2 seconds [30]. The amount of nitrites and nitrites in the diet vary substantially. However, in combination with other prooxidants in the digesta they can be involved in free radical formation and lipid peroxidation.
Heavy metals
Agricultural uses of phosphate fertilizers and sewage sludge and industrial uses of cadmium have been identified as a major cause of widespread dispersion of the metal at trace levels into animal feed and human foodstuffs. It is well known that heavy metals can cause oxidative stress and stimulate lipid peroxidation [31]. For example, cadmium increased lipid peroxidation in liver, kidney, and testes of rats and reduced metallothionein and total sulfhydryl in liver and kidney [32]. In renal tubular epithelial cells of rats significant decrease in activities of GSH, GSH-Px, SOD and increased malondialdehyde (MDA) formation were observed as a result of the treatment with lead and cadmium [33]. It is believed that lead can alter certain membrane bound enzymes and may cause oxidative stress. For example, exposure of HepG2 cells to lead ions decreased cell viability and stimulated lipid peroxidation of cell membranes decreasing the fluidity in the polar surface of cell membranes [34]. Levels of lipid peroxidation products such as MDA, conjugated diene and hydroperoxide were increased in liver, lung and kidney of lead-treated rats. Consumption of lead in drinking water by rats imposed oxidative stress increasing lipid peroxidation in peripheral blood mononuclear cells and liver [35]. Administration of exogenous antioxidants in the lead treated animals significantly reduced the prooxidant effect of the toxicant [36]. Mercury is also a strong prooxidant able to increase lipid peroxidation and decrease GSH content in liver of Swiss albino mice. In particular, mercury treatment enhanced lipid peroxidation in kidney, testis and epididymus of rats [37]. Heavy metal concentrations in major feed and food sources are quite low; however, in combination with other prooxidants they potentially can be involved in generation of free radicals and cause oxidative stress in the GIT.
Persistent organic pollutants
Persistent organic pollutants (POPs) comprise a class of chemicals that are among the most insidiously dangerous compounds and it includes many organochlorine pesticides. Examples of persistent organic pollutants found in food and feed include dioxins, polychlorinated biphenyls (PCBs), polybrominated diphenyl ethers, and some pesticide chemicals [38]. Contaminating/accumulating levels of organochlorine pesticides in extractable fats from a basal diet, eggs and seven tissues and excreta of laying hens that were kept in a general poultry farm of Japan were examined [39]. Dieldrin and dichlorodiphenyltrichloroethane (DDT) were shown to contaminate the dietary fats at the level of 10-70 ppb. In 1989-1994, the U.S. Food and Drug Administration (FDA) analyzed 545 samples of mixed feed rations and found that only 88 samples (16.1%) did not contain detectable pesticide residues [40]. Malathion, chlorpyrifosmethyl, diazinon, chlorpyrifos, and pirimiphos-methyl were the most commonly detected pesticides comprising 93.4% of all pesticide residues detected. Their median values in samples containing quantifiable levels ranged from 0.014 to 0.098 ppm.
Animal products like milk and meat are often found to be contaminated with residues of persistent pesticides and other toxic substances. The major entry source of these compounds to animal body is the contaminated feed and fodder. The frequency of occurrence and contamination levels of organochlorinated pesticides (OCP) residues in different kinds of animal concentrate feed and straw samples collected from Bundelkhand region of India were determined [41]. Out of 533 total samples, 301 i.e. 56.5% samples were positive containing residues of different OCPs and in the case of DDT complex, i.e. DDD, DDE and DDT, the concentration ranged between 0.016 and 0.118 mg/kg. It has been shown that organochlorines can cause oxidative stress. For example, the adverse effect of organochlorine pesticide methoxychlor on the male reproductive system was shown to be due to induction of oxidative stress in testis [42,43]. Similarly, a pesticide hexachlorocyclohexane compromised antioxidant defence and induced oxidative stress in rat cerebral hemisphere[44]. Even if the levels of heavy metals are in the accepted limits, they are still able to participate in lipid peroxidation. Therefore, persistent organic pollutants represent an important health hazard for animals and they also can be involved in promotion of lipid peroxidation in the gut.
Mycotoxins
At least 25% of world’s grain production is contaminated with mycotoxins, which are a worldwide problem [45]. Indeed, mycotoxins are the major feed- and food-derived stressors [46]. Recently, the occurrence of ochratoxin A (OTA) in complete poultry feeds (n = 80) and poultry feed ingredients (n=286) from Pakistan has been evaluated [47]. Contamination frequency and mean OTA levels were 31% and 51 μg/kg in feed ingredients, and the corresponding values for complete feeds were 38% and 75 μg/kg. In an Argentinian study of mycotoxins in poultry feedstuff, forty-four out of 49 samples (90%) were contaminated with deoxynivalenol (DON; median 222 ppb) and OTA (median 5 ppb). In addition, 44 out of 49 samples were contaminated with aflatoxins (median 2.7 ppb), 42 samples (86%) with zearalenone (ZEA; median 50 ppb), and 38 samples (78%) with T2-toxin (median 50 ppb; [48]). A total of 55 feed ingredients as well as 76 complete swine feeds were randomly collected from 15 swine farms located in the Beijing region of China from July to August 2011 and mycotoxins were analysed [49]. It is important to note that DON and ZEA were the most prevalent mycotoxins found. DON was detected at percentages of 93, 92, 54, 100 and 97% with a mean level of 1.01, 0.44, 0.05, and 0.65 ppm in the samples of corn, wheat bran, soybean meal, and complete feeds, respectively. The detected percentages of ZEA were 100, 100, 54 and 100 with mean levels of 109.1, 14.9, 9.2 and 58.9 ppb in the same samples. In another study from China, of the 420 analysed feedstuff samples, the incidence of T-2, ZEN and fumonisin B1 (FB₁) was 79.5%, 85.2% and 96.1%, respectively and detected concentrations ranged from 10-735, 35- 1478 and 20-6568 μg/kg, respectively [50]. Indeed, during an 8-year period, 17316 samples of feed and feed raw materials from all over the world were analysed for contamination with aflatoxins, OTA, ZEA, DON and fumonisins. Overall, 72% of the samples tested positive for at least one mycotoxin and 38% were found to be co-contaminated [51]. Therefore, the chances of getting mycotoxins in pig/poultry diet are very high.
It is necessary to mention that OTA altered both barrier and absorption function of the intestinal epithelium causing intestinal injuries, including inflammation and diarrhea [52]. It is interesting that inhibition of protein synthesis and induction of apoptosis are the main mechanisms of DON toxicity in intestinal cells [53]. In general, main mycotoxin contaminants of the food including aflatoxin B1 (AFB1), fumonisins, T-2 toxin, DON, ZEA and OTA are shown to compromise antioxidant system and stimulate lipid peroxidation in vivo and in vitro [54,55,46]. Mycotoxins are considered to be unavoidable contaminants of the most food and feed ingredients and they are potent prooxidants able to affect a range of genes [56]. Therefore, even in comparatively low concentrations (lower than the officially allowed limits) they still represent an important source of free radical generation in the GIT [57,61].
Immune system
The immune system is considered to be an important source of ROS in the human body [54] and intestinal immunity is not an exception. Indeed, the intestinal tract is considered to represent the largest immune organ of the human body responding to the challenge of bacteria or food antigens by production of ROS [62]. Mucosal surfaces covered by a layer of epithelial cells represent the most critical interface between the organism and its environment since the mucosal interstitia of the intestine is continuously exposed to large amounts of dietary and microbial antigens. Therefore, epithelial cells engage in cross talk with luminal bacteria and their products and produce mediators and signals that are key components of host innate and acquired mucosal immunity [63]. The mucosal immune system is a first line of defence against foreign antigens, including microbial and dietary antigens and under normal circumstances it employs tightly regulated dynamic mucosal intra- and internets consisting of inductive (e.g. Peyer’s patch) and effector (e.g. intestinal lamina propria) tissues and maintains an appropriate immunological homeostasis between the host and mucosal environments [64]. Hence, the mucosal immune system has evolved efficient mechanisms to distinguish potentially pathogenic from non-pathological antigens. For example, the mucosal immune compartment must be able to choose the appropriate effector function (e.g., tolerance vs. clearance) necessary to deal with each encountered antigen whether it is innocuous or pathogenic in nature [65]. However, abrogation of these mucosal defence mechanisms may alter immunological homeostasis in the gastrointestinal tract and induce pathological changes including chronic active inflammation, mucosal atrophy andtissue injuries [66]. It is important to stress that under inflammatory conditions in the intestine the maintenance of the epithelial barrier could be broken.
It is necessary to underline that the nutrient requirement to maintain a highly active immune system in the digestive tract could be quite high. In fact, a different source of injury to the intestinal mucosa (nutritional, infectious or allergic) act via a common mechanism of cell-mediated immune damage and nutrient repletion is required for restoration of immune function [67]. In particular, the immunomodulating properties of natural antioxidants [54] could be of great advantage for the intestinal immunity. Furthermore, intestinal epithelium can modulate the level of immune activity in the mucosal immune system according to the environment of the intestinal lumen [68]. Data presented above indicate that intestinal immune system can generate free radicals in response to various antigens including microbes and some food allergens.
Combinations of prooxidants in the gut and their detrimental effects
Physicochemical environment of the gastrointestinal tract depends on many factors with diet, bacterial metabolites andbody secretion being major determinants [68]. There is a delicate balance between the environment of the lumen and epithelial cell functionality and dietary factors are responsible for gene expression in the intestine and its adaptation. In this regard, oxidative stress could cause changes in this balance affecting absorption of nutrients. Even if each of those lipid peroxidation promoters is present at a very low concentration, their combination could be much more powerful. For example, as mentioned above lipid hydroperoxides can produce peroxyl radicals in presence of iron or copper ions and once chain reaction of lipid peroxidation started many other food-derived PUFAs can be oxidized. It was calculated that pH and temperature,as well as presence of oxygen in stomach could be favourable for lipid peroxidation [69].
Data provided above indicate that average pig or poultry diet contains a range of various prooxidants. They include oxidized polyunsaturated fatty acids, nitrites, nitrates, heavy metals,mycotoxins, persistent organic pollutants, etc. In many cases those contaminants are found in the feed in low or very low concentrations, however, their various combinations in the fee could be an important source of free radical production in the gut.
Antioxidant Defences in the GIT
An effective antioxidant protection in the gastrointestinal tract is needed to maintain gut health and this protection is based on food derived antioxidants [31].Vitamin E
Vitamin E is main biological chain-breaking antioxidant, located in the biological membranes of various tissues. In food and feed ingredients vitamin E can be found in the form of 4 tocopherols and 4 tocotrienols. It is possible that the gut is a special place for γ-tocopherol and tocotrienols to play their antioxidant role. Alphatocopherol is not stable and easily oxidized during food processing and therefore commercial vitamin E preparations are represented by the stable esterified form of vitamin E or as a mixture of tocopherols. For farm animals and poultry premixes are the main source of vitamin E and its level in feed ingredients are often not taken into account during feed formulation [54].
The main reason for vitamin E dietary supplementation for poultry and farm animals is to maintain their optimal health and high productive and reproductive performances. This includes positive effects on male and female reproduction, immunocompetence,effective growth and development, high quality of eggs and meatas well as decreased negative consequences of various stresses [54,31,70,74]. Extensive research and wide commercial application for a number of years clearly showed essentiality of vitamin E in animal/poultry nutrition. Recently it has been shown that vitamin E recycling in the cell is the key for its antioxidant activity. Ascorbic acid, selenium, vitamins B1 and B2 are important elements of vitamin E recycling. Therefore, if recycling is effective, even a low vitamin E concentration, for example in the embryonic brain, can prevent lipid peroxidation in vivo [54,70]. There is a range of anti-stress premixes with increased vitamin E content but their efficacy is variable. After 90 years of extensive research in the field of vitamin E we greatly appreciate its unique role in biological systems, in maintaining growth, development and general health of humans and animals. High concentrations of vitamin E were present in the mucosa of the duodenum and jejunum, with a trend to lower levels in the ileum and ceca, and significantly less in the colon of the chicken [75]. The different isoforms of vitamin E were absorbed from the digesta by the mucosa without any major selectivity. However, the liver was greatly enriched with alpha-tocopherol over the other isoforms, indicating a high degree of discrimination by this tissue. Clearly, vitamin E can be considered as a main contributor to the antioxidant potential of the digesta [31,74].
It should be also mentioned that beyond a direct antioxidant activity, vitamin E is involved in modulation of enzyme transcription and/or activity by interacting with genes involved in oxidative stress, proliferation, inflammation and apoptosis [76]. In fact, such genes include SOD, NO synthase, cyclooxygenase-2, NAPDH oxidase, NF- κB, phospholipase A2, protein phosphatase 2A, 5-lipooxygenase, activator protein-1, cytochrome P450, BCL2-like 1 and a lot of other genes. However, it is not clear at present if all aforementioned genes are affected by vitamin E in the gut.
Coenzyme Q
Coenzyme Q (CoQ), known also as ubiquinone, was discovered in 1957. The name ubiquinone is related to its ‘ubiquitous” presence in all cells and the name coenzyme Q reflects the chemical structure of the compound containing one quinone group and 10 isoprenyl units. Coenzyme Q10 (CoQ10) exists both in an oxidised and a reduced form, ubiquinone and ubiquinol, respectively [77]. In general, dietary supplementation of CoQ does not affect the endogenous synthesis of CoQ in tissues. However, oxidative stress (physical exercise, thyroid hormone treatment, cold adaptation, vitamin A deficiency, etc.) is associated with increased CoQ synthesis reflecting a cellular adaptation [78]. Therefore, CoQ synthesis is considered to be an adaptive mechanism in response to stress conditions when other antioxidants are depleted. For example, in vitamin E and Se deficient rats CoQ concentration elevated and CoQ-dependent reductase system is activated [79].
Antioxidant properties of CoQ are directly related to the protection in the gastrointestinal tract. For example, in rats treated per os with sodium nitrite increases TBARS in small intestinal mucosa and liver were observed. Pre-treatment of nitrite-poisoned rats with CoQ10 mitigated lipid peroxidation and increased total antioxidant status in animal blood [80]. The protective effect of administered CoQ10 against small intestinal damage caused by ischemia reperfusion was also shown [81]. In rat intestine, administration of CoQ10 normalised a sharp gamma-irradiation-induced inhibition of transformation of phosphatidylcholine from phosphatidylethanolamine [82]. Compared to paired non-inflamed mucosa, concentration of CoQ10 was significantly decreased in inflamed mucosa [83]. The decreased antioxidant defences may severely compromise the inflamed mucosa, rendering it more susceptible to oxidative tissue damage, hindering recovery of the mucosa and return of epithelial cell layer integrity. Therefore, antioxidant and other regulating functions of CoQ10 could be extremely important in the GIT.
Carotenoids
Carotenoids comprise a family of more than 750 compounds responsible for a variety of bright colours in nature [84]. It is well appreciated that carotenoids are important elements of the antioxidant system, possessing antioxidant activities and participating directly or indirectly (for example, by recycling vitamin E or regulating expression of various genes) in antioxidant defences. Recently, an important role of canthaxanthin with a special emphasis to carotenoid antioxidant activities in breeder nutrition has been described [85,86]. Biological functions of these natural pigments in relation to animals or humans are not well defined but their antioxidant properties seem to be of major importance. In mixture with other antioxidants they could be much more effective than on their own, and the GIT could be a major place for these compounds to exert their activity. Furthermore, carotenoids can induce the expression of genes related to antioxidant defences. For-example, recently it has been shown that lutein modulates the expression of a range of genes related to oxygentransporters and decreases DNA damage and oxidative stress in miceCarotenoid assimilation from the diet varies significantly depending on many various conditions; however, it seems likely that a substantial proportion of ingested carotenoids could be found in all segments of the digestive tract. In particular the chicken mucosa of the duodenum and jejunum contained the highest concentrations of carotenoids, with much lower levels in the ileum and colon [75].Therefore, in combination with other dietary antioxidants carotenoids could promote antioxidant defence in the gut. Furthermore carotenoid activities related to the promotion of cell differentiation, regulation of cell proliferation and intracellular communication via gap junctions, as well as regulation of the detoxifying enzymes and enhancement of immune system [54,85,86] could also be of great importance in the gut.
Ascorbic acid (AA)
Vitamin C is referred to as L-ascorbic acid and its two-electron reduction product dehydro-L-ascorbic acid. Most animal species synthesize AA from glucose, but human subjects are not able to synthesize it. Therefore, AA is an essential dietary component playing an important role in many physiological processes and it is a hydrophilic antioxidant functioning in an aqueous environment and possessing high free-radical-scavenging activity. It can participate in vitamin E recycling thus maintaining efficient antioxidant defence [31]. Due to its high reducing potential, in combination with iron ions AA can be a prooxidant. However, it is believed that in physiological conditions and in the GIT ascorbic acid performs mainly its antioxidant functions. In fact, ascorbic acid inhibits chemical synthesis of nitrosamines (animal carcinogens) in the gastric contents and there are suggestions that intakes of ascorbic acid much higher than RDA may reduce the risk of such diseases as heart disease and cancer [88]. Ascorbic acid is synthesised in farm animals and poultry, but in stress conditions, additional dietary vitamin C supplementation or its provision with drinking water is shown to be helpful.
Glutathione
Glutathione (GSH) is the most abundant non-protein thiol in mammalian cells, and is considered to be an active antioxidant in biological systems providing cells with their reducing milieu. It is well known that GSH can be synthesised in the human body and it is abundantly distributed in the mucosal cells of the GIT in man and its highest concentration is found in the duodenum [89]. Cellular GSH plays a key role in many biological processes: the synthesis of DNA and proteins, including cell growth and proliferation, regulation of programmed cell death, immune regulation, the transport of amino acids, xenobiotic metabolism, redox-sensitive signal transduction [90,91]. Furthermore, GSH thiolic group can react directly with H2O2, superoxide anion, hydroxyl radicals, alkoxyl radicals, hydroperoxides [92]. Furthermore, in stress conditions GSH prevents the loss of protein thiols and vitamin E and plays an important role as a key modulator of cell signalling [93]. Animals and human are able to synthesise glutathione. In addition to body synthetic activity feed also provide GSH. In general, it is difficult to overestimate a protective role of GSH in the gut [94,97].
Polyphenols/flavonoids
Natural polyphenols comprises a big group of compounds with flavonoids, low molecular weight polyphenolic substances based on the flavan nucleus, being the most studied ones. They are widespread in nature, occurring in all plant families. The list of flavonoids substantially increased in the last decades accounting for over than 8000 individual compounds [98]. It seems likely that the gut is the major place of antioxidant action of polyphenols [73]. Indeed, reduction of oxidative damage, modulation of colonic flora and variation in gene expression are involved in the modulation of intestinal function by polyphenols. For example, to study the molecular effects of wine polyphenols at the gene level, the microarray technology was used: rats were treated with 50 mg/kg wine polyphenols for 14 days, mixed in the diet. It was shown that two major regulatory pathways were down-regulated in the colon mucosa of polyphenols treated rats: inflammatory response and steroid metabolism [99]. Since flavonoids are consumed in concentrations usually much higher than other antioxidant compounds, their protective effect during digestion is of great importance. For example, it has been shown that flavonoids not only prevented an accumulation of peroxidized lipids but also could switch prooxidant properties of heme-proteins to antioxidant ones [69]. Dietary polyphenols can also modulate invivo oxidative damage in the gastrointestinal tract of rodents [100] supporting the hypothesis that dietary polyphenols might have both a protective and a therapeutic potential in oxidative damage-related pathologies. Indeed, the antioxidant-prooxidant balance (redox status) in various parts of the intestine would ultimately depend on the level of antioxidants and prooxidants provided with the diet and released by cells themselves as well as on the level of absorption of both antioxidants and prooxidants. In a model system mimicking stomach conditions it was shown that both lipid peroxidation and co-oxidation of vitamin E and beta-carotene were inhibited at pH 3.0 by red wine polyphenols [101].
Redox signaling in gut inflammation is complex and poorly understood. However, it is generally accepted that homeostatic control of the intestinal epithelial redox environment is central for nutrient digestion and absorption, stem cell proliferation, apical enterocyte apoptosis, and immune response [102]. Indeed, polyphenols may play a role on intestinal mucosa integrity, inflammation and permeability [103]. For example, wine phenolics were able to prevent or delay the progression of intestinal diseases characterized by oxidative stress and inflammation, acting as both free radical scavengers and modulators of specific inflammation-related genes involved in cellular redox signaling [104]. They exert their effects by modulating cell signaling pathways, mainly activated in response to oxidative and inflammatory stimuli, and Nrf2 and NF-κB are the principal downstream effectors [105]. It is possible to suggest that there is a biological reason for some antioxidants not to be absorbed completely and in that way providingantioxidant protection in lower parts of the intestine. Comparatively low bioavailability and antioxidant potential of various flavonoids could be beneficial for the human/animals providing antioxidant protection in various parts of the digestive tract, including the large intestine where levels of other antioxidants would be quite low.
Synthetic antioxidants
Antioxidants in feed may be of endogenous origin or may be added externally to preserve their lipid components from peroxidation. Synthetic antioxidants such as ethoxyquin and its blends with other antioxidants such as propyl gallate [106,109] are used to stabilise fatin poultry and pig diets and they play an important role in antioxidant defences of the gut.
Specific place for Se-dependent enzymes in antioxidant defence of the GIT
Food derived antioxidant enzymes would be inactivated during thermal food processing. However, the GIT contains internallyoriginated antioxidant enzymes SOD, GSH-Px and CAT and they represent an important mechanism of the enterocyte defence from oxidative damage. A specific gastrointestinal GSH-Px (GI-GSH-Px) has been described in 1993 [110]. GI-GSH-Px activity was present in both the villus and crypt regions of rat mucosal epithelium and its activity nearly equalled that of classical GSH-Px throughout the small intestine and colorectal segments [111]. GI-GSH-Px could be considered to be a barrier against hydroperoxide resorption [112,113]. Furthermore, in the gastrointestinal tract there are at least three more selenoproteins including plasma GSH-Px, selenoprotein P and thioredoxin reductase [114].
Glutathione and glutathione–dependent enzymes contribute significantly towards intestinal antioxidant defences. In fact, an important peroxide detoxification pathway in the intestine is based on the GSH redox system [115]. In this system GSH-Px reduces peroxides at the expense of GSH oxidation. Oxidised glutathione is reduced back to the active form by glutathione reductase utilizing reducing potential of NADPH which is produced in the pentose phosphate pathway.
Role of vitagenes in the gut defence
To adapt to environmental changes and survive different types of injuries, eukaryotic cells have evolved networks of different responses which detect and control diverse forms of stress. Recently, a vitagene concept has been developed. In fact, the term “vitagene”was introduced in 1998 by Rattan [116] who considered them as a range of genes to be involved in regulation of various protective mechanisms in stress conditions. Later a vitagene concept has been further developed by Calabrese and colleagues [117-122] with major emphasis to prosurvival mechanisms controlled by vitagene network. Furthermore, possible roles of vitagenes in protection of chickens against various stresses has been reviewed recently [123-126]. In accordance with Calabrese et al. the vitagene family includes genes that are strictly involved in preserving cellular homeostasis during stress conditions [118,119]. In fact, the vitagene family includes heatshock proteins (Hsps), such as haeme oxygenase-1 (Hsp32, HO-1), Hsp60 and Hsp70, the thioredoxins (Trx)/ thioredoxins reductase(TR) system and sirtuins. It seems reasonable to extend the list of potential candidates to vitagene family. In particular, SOD, a major inducible enzyme of the first level of antioxidant defence, can meet selecting criteria to be included into the vitagene family. The products of the mentioned genes are responsible for detection and control of diverse forms of stress and cell injuries. The cooperative mechanisms of the vitagene network are reviewed in recently published comprehensive reviews [119,121,122,127] with a major conclusion indicating an essential regulatory role of vitagene network in cell and whole organism adaptation to various stresses.
Recently, new mechanisms of such adaptive defences of the gastrointestinal mucosa at the intracellular level have been characterised. One of these responses, known as the heat shockresponse is considered to be a universal fundamental mechanism necessary for cell survival under a variety of unfavourable conditions [128]. As mentioned above, intestinal cells are challenged with a great variety of potentially toxic compounds and their protection is a vital part of the strategy to maintain human health. In mammalian cells, the induction of the heat shock response requires the activation and translocation to the nucleus of one or more heat shock transcription factors, which control the expression of a specific set of genesencoding cytoprotective heat shock proteins [128]. Indeed, Hsps have a broad range of functions related to their major role in cellular homeostasis and protect cells against apoptosis and cell death. The physiological expression of cytoprotective Hsp27 in the gut was investigated in eighteen 7-wk-old pigs [129]. Indeed, Hsp27 was expressed in all the samples from ileum and colon and the expression was most intensive in the apical intestinal epitheliums in close contact with luminal contents and lighter in crypt cells. Furthermore, the ileal Peyer’s patches showed a strong expression of Hsp27, which was highly correlated with Hsp27 expression in the ileal epithelial cells. The expression of Hsp27, heat shock cognate 70 (HSC 70), Hsp70 and Hsp90 along the GIT of young pigs and the effect of weaning on this expression were studied [130]. Pigs were weaned at 28 or 21 d and slaughtered at various times post-weaning. All Hsps were expressedin the GIT segments studied before and after weaning. However, there was a site specificity in Hsp expression in the gut. For example, the expression of Hsp27 and Hsp70 was increased in the stomach and duodenum between 6 and 12 h post-weaning and between 24 and 48 h in the mid-jejunum, ileum and colon. At the same time, their expressions were transiently decreased in the ileum. Expression of Hsp90 increased in the stomach and jejunum but decreased in the duodenum, ileum and colon. Similar results were obtained at both ages of weaning. Indeed, in normal porcine GI tract Hsp expression is gut region- and cell type-specific in response to dietary components, microbes, and microbial metabolites to which the mucosa surface is exposed [131].
Therefore, Hsps function as molecular chaperones in regulating cellular homeostasis and promoting survival. However, if the stress is too high, a signal that leads to programmed cell death, apoptosis, is activated, thereby providing a finely tuned balance between survival and death [132]. In addition to extracellular stimuli, several stressful conditions induce Hsps during normal cellular growth and development. In particular, the Hsp family is activated under oxidative stress and provides an important protection against protein denaturation and modifications by capping and refolding, or drives damaged proteins into appropriate proteolytic pathways [133]. In fact, Hsps have been assigned to multiple subcellular sites and implicated in multiple functions ranging from stress response, intracellular trafficking, antigen processing, control of cell proliferation, differentiation, and tumorigenesis [134]. Therefore, in response to environmental or physiological stresses cells increase synthesis of Hsp [135]. It has been suggested that the conserved heat shock protein Hsp33 functions as a potent molecular chaperone with a highly sophisticated regulation. In fact, at the transcriptional level, the Hsp33 gene is under heat shock control; at the posttranslational level, the Hsp33 protein is under oxidative stress control [136].
Therefore redox-regulated chaperone activity of Hsp33 specifically protects proteins and cells from the detrimental effects of reactive oxygen species.
ROS-mediated damage has been implicated in the pathophysiology of the gastrointestinal mucosa and Hsps are suggested to play an important role in cytoprotection against oxidative stress-induced injury [137]. For example, the mammalian intestinal epithelial cells respond to heat stress by producing heat shock proteins that provide protection in stress conditions, which would otherwise lead to cell damage or death. The protective effects of Hsps are seen in heat stress, infection, and inflammation [138]. Similarly, glucocorticoid protection of rat intestinal cells against oxidant-induced stress was mediated by Hsp72 [139].
The molecular mechanisms of heat shock response-induced cytoprotection are beyond this review. However, they involve inhibition of proinflammatory cytokine production and induction of cellular proliferation for restitution of the damaged epithelium [138]. It is interesting to note that Hsp72, the stress-inducible form of Hsp70, was detected in samples from rat distal colon, proximal colon, and terminal ileum, but was not found in proximal small bowel or other organs (liver, kidney, spleen, heart, and brain) of unstressed animals [140]. Hsps play an important role in gastric mucosal defence under conditions of stress. For example, exposure of rats to restraint and water-immersion stress caused rapid Hsp70 mRNA expression and Hsp70 accumulation in gastric mucosa and the extent of Hsp70 induction inversely correlated to the severity of mucosal damage [141]. Therefore, Hsp70 is involved in repair of partially damaged proteins and substantially contributes to protection of the gastrointestinal mucosa against various necrotising factors [135].
It is known that HO-1, known as Hsp32, can be induced by various stresses. Indeed, HO-1 induction and the maintenance of its appropriate activity is critical in protecting the intestinal epithelial cells from oxidative injury [142]. It is interesting that in the aforementioned experiment HO-1 was markedly induced following LPS treatment in the mucosal epithelial cells in the upper intestine (duodenum and jejunum) but not in the lower intestine (ileum and colon). It seems likely, that there is a delicate interaction between Hsps and other antioxidant defence mechanisms to maintain mucosal integrity and repair of acute mucosal damage.
Thioredoxin and TR are considered to be also important members of vitagene family. Their role in the gut is not clear at present, but, clearly they participate in maintenance of the redox balance in the gut. The presence of a full complement of Trx/TR proteins in stomach, duodenum, jejunum, ileum and colon suggests their function in antioxidant defense and redox regulation in the intestinal tract [143]. It is important to note that Trx expression is particularly high in the small intestinal mucosa and colon [144], while TR expression in small intestine is substantially higher than in colon. All three thioredoxin reductases are expressed in the intestine at least at the mRNA level, which is relatively unaffected by marginal selenium deficiency and in the case of TR2 and TR3 rather increased when selenium becomes limited [145]. Furthermore, the selenoproteins GPx2, TR2 and TR3 in the gut are regulated by the Wnt pathway [146]. It has been also shown that unstimulated lamina propria T lymphocytes exhibited high expression of Trx which involved in the regulation of intracellular redox homeostasis in these cells [147]. The authors suggested that Trx may play a key role in the specialized intestinal microenvironment in amplifying immediate immune responses. In fact, membrane-bound Trx converts human β-defensin 1 to a potent antimicrobial peptide in vivo [148]. Takaishi et al. identified rat Trx as a growth-promoting factor for intestinal epithelial cells [149]. Higashikubo et al. showed that cellular oxidative shock caused an increase in the activity of thioredoxin, which is involved in the defense mechanism against oxidative stress [150]. In particular, H2O2 was cytotoxic to the small intestine epithelial cell line, IEC-6 and the glutathione S-transferase and thioredoxin reductase activities and SH content decreased dosedependently with H2O2, while thioredoxin activity increased at low H2O2 concentrations.
Critical periods of the gut development: chick placement
Chick viability is an important factor in determining profitability and, from fertilisation to placement at the broiler farm, factors such as egg quality, egg storage conditions, incubation conditions and post-hatch environment will all affect chick quality [151]. It is well appreciated that time between chick hatch and placement is stressful due to dehydration and yolk sac reserve depletion. Indeed, if we put together hatching time inside the hatcher, time of chick processing and transportation, and finally, placement at the farm, it could take up to 36-48 h before a newly hatched chick has access to feed and water and during this time body weight decreases quickly [152]. It has been shown that in the hatching chick, the small intestine matures in a manner similar to neonatal mammals, with specific ontogenetic timetables in the different small intestinal segments; however, the most dramatic changes occur within the first 24 h post-hatch [153]. There is an inverse relationship between duration of post-hatching olding time and subsequent chick performance [154,155]. Therefore, immediate access to feed and water can increase body weight of the growing chick at 3 weeks of age [156] or at market age of broilers [157]. It should be mentioned that there is the hatch window (24-36 hours) or the spread between late and early hatchers which depends on the homogeneity/heterogeneity of the incubating eggs including egg size and breeder age. A spread in the hatching period will increase the numbers of chicks sitting extra hours in stressful conditions of the hatcher without food or water. In fact, hatching process places a stress upon the emerging chick and natural antioxidants (vitamin E and carotenoids) have evolved to reach the maximum concentration in the liver to protect the unsaturated lipid in the tissues and so limit lipid peroxidation [158,159]. Furthermore, any delay in accessing food [160,161] and/or water intake after hatching as well as hatchery treatments such as vaccination, sexing and transport to the farm can result in additional stress [162]. Indeed, extended time in the hatcher (36 h) was associated with decreasing vitamin E coenzyme Q concentrations in chicken tissues [163]. Given the relatively high temperature and humidity in the hatcher, it is easy to make the argument that the chick may be under chronic oxidative stress during this holding time. Therefore, antioxidant protection at hatching time is considered to be an important determinant of chick viability during first post-hatch days [54,164-166]. During chick embryo development there is an antioxidant/prooxidant balance in the tissues which supports normal embryonic development and post Chick viability is an important factor in determining profitability and, from fertilisation to placement at the broiler farm, factors such as egg quality, egg storage conditions, incubation conditions and post-hatch environment will all affect chick quality [151]. It is well appreciated that time between chick hatch and placement is stressful due to dehydration and yolk sac reserve depletion. Indeed, if we put together hatching time inside the hatcher, time of chick processing and transportation, and finally, placement at the farm, it could take up to 36-48 h before a newly hatched chick has access to feed and water and during this time body weight decreases quickly [152]. It has been shown that in the hatching chick, the small intestine matures in a manner similar to neonatal mammals, with specific ontogenetic timetables in the different small intestinal segments; however, the most dramatic changes occur within the first 24 h post-hatch [153]. There is an inverse relationship between duration of post-hatching holding time and subsequent chick performance [154,155]. Therefore, immediate access to feed and water can increase body weight of the growing chick at 3 weeks of age [156] or at market age of broilers [157]. It should be mentioned that there is the hatch window (24-36 hours) or the spread between late and early hatchers which depends on the homogeneity/heterogeneity of the incubating eggs including egg size and breeder age. A spread in the hatching period will increase the numbers of chicks sitting extra hours in stressful conditions of the hatcher without food or water. In fact, hatching process places a stress upon the emerging chick and natural antioxidants (vitamin E and carotenoids) have evolved to reach the maximum concentration in the liver to protect the unsaturated lipid in the tissues and so limit lipid peroxidation [158,159]. Furthermore, any delay in accessing food [160,161] and/or water intake after hatching as well as hatchery treatments such as vaccination, sexing and transport to the farm can result in additional stress [162]. Indeed, extended time in the hatcher (36 h) was associated with decreasing vitamin E and coenzyme Q concentrations in chicken tissues [163]. Given the relatively high temperature and humidity in the hatcher, it is easy to make the argument that the chick may be under chronic oxidative stress during this holding time. Therefore, antioxidant protection at hatching time is considered to be an important determinant of chick viability during first post-hatch days [54,164-166]. During chick embryo development there is an antioxidant/prooxidant balance in the tissues which supports normal embryonic development and posthatch chick viability. It has been suggested that an accumulation of the natural antioxidants like vitamins A, E and carotenoids as well as an increase in GSH-Px activity in the embryonic liver may have an adaptive significance, evolving to protect unsaturated lipids against peroxidation during the stress imposed by hatching [54].
The antioxidant system of the chicken embryo and newly hatched chick has been studied extensively [54]. It was shown that it includes fat-soluble antioxidants (vitamin E and carotenoids) originating from the maternal diet [159], as well as water-soluble antioxidants (ascorbic acid, glutathione, and uric acid) and antioxidant enzymes (superoxide dismutase, glutathione peroxidase and catalase) which are synthesised during embryonic development [167,168]. Previously, a dramatic decrease in vitamin E concentrations in the chicken liver for the first 10 days of post-hatch development has been shown for chickens, turkey, duck and goose [165]. An increase in dietary vitamin E supplementation slowed down this process but did not change the trend [54,166]. Therefore, one of the most impressive features of vitamin E metabolism in avian embryonic tissues is an abrupt decrease in the concentration of this vitamin over the first two weeks of postnatal development. As indicated previously, the liver accumulates vitamin E during embryonic development to supply chickens with this vitamin in the first days of life after hatch [159]. This reserve of vitamin E is used by chickens during the first 2 weeks post-hatch. During this period vitamin E concentration in the liver decreased by 10 times in chickens, goslings and ducklings [54] and more than 50 times in turkeys [169]. Marusich et al. suggested that the low levels of vitamin E in the liver of turkey poults resulted from the inefficient intestinal absorption of vitamin E [170], which can be explained as a result of low pancreatic lipase activity [171] and restricted bile production [172] as well as of greater (compared to chicken) production and excretion of tocopheryl glucuronides [173]. In general, the capacity for fat and probably fat-soluble vitamin E absorption is incompletely developed in the newly-hatched chick, but it matures rapidly in the first week of life [172,174]. On the other hand, during this period of development, tissues with incompletely developed antioxidant regulation require an effective protection against lipid peroxidation, which is afforded mainly by vitamin E. These data also confirmed the biological importance of a very high vitamin E concentration in the embryonic liver at hatching time.
Postnatal nutritional exposures are considered to be critical for the developmental maturation of many organ systems and optimalphysiological functions. There is a growing body of evidence indicating that environmental exposures including nutritional exposures during these critical and sensitive periods of life can cause permanent changes in many physiological processes, which is known as “programing” [175]. In this regard, research data are accumulating to support the hypothesis that the vitamin E status of turkey poults and probably chickens may be inadequate during the first 3 weeks after hatching [176]. A variety of approaches aimed at improving the vitamin E status of turkey poults have, in fact, been investigated; these have included dietary supplementation of the poults with high levels of α-tocopherol [54,177], bile salts [169] and fat [178], as well as vitamin E injection [179] and alterations in provision of n-6 and n-3 polyunsaturated fatty acids [177]. When d-α-tocopherol was added in the drinking water, there was a temporary increase of α-tocopherol in tissues and a decreased susceptibility of red blood cells to hemolysis [180]. Moreover, day-old chickens were treated with 3.25 mg vitamin E/bird/day per os, via the drinking water, for two weeks. The vitamin E content of both the liver and the blood plasma was significantly higher in the treated chickens than in the untreated controls [181]. It seems likely, that provision of vitamin E and other fat-soluble vitamins (A and D3) with water at time of chicken placement can solve the problem of their low availability for newly hatched chicks. Such a supplementation helps chickens overcome stress of placement and has positive effect on chicken growth and development. The new concept of fighting stresses was based on an idea that supplying birdswith various antioxidants via the drinking water could help them deal with stress conditions more effectively. Indeed, it was proven that inclusion of vitagene-regulating compounds (vitamin E, Vitamin A, in combination with carnitine, betaine and other compounds) in water could be effective in fighting various stresses [123-125,182]. This helps at chick placement, when the antioxidant system is crucial for the digestive and immune system development [182]. In particular, it was proven that inclusion of an anti-stress composition (PerforMax) at time of chicken placement into the drinking water at the University trial improved chicken growth and feed conversion ratio (FCR) [183,184]. Using the same anti-stress composition in commercial conditions improved FCR during a 39-day broiler growth trial. At the end of the trial, the improvement in FCR due to the anti-stress composition during the first three days post-hatch as well as before and after vaccination was highly significant [126,185]. In addition,it was shown that the anti-stress composition had an immunemodulating effect in broilers [183], growing ducklings [186] and could be successfully used to prevent immunosuppression [187,188]. Improvement of the antioxidant system via supplying the antioxidant composition via the drinking water could help deal with various mycotoxins in feed, including DON [189,190], OTA [191,192], and T-2 toxin [193,194]. Furthermore, such a technology could also help fighting heat stress [183,195,196].
Our previous investigations indicate that low quality neonatal nutrition resulted in long-term impairment in the capacity to assimilate dietary antioxidants. In fact, birds that had experienced the relatively low-quality diets during their early growth had half the levels of lipophilic antioxidants (vitamins A, E and carotenoids) as adults than birds reared on the standard-quality diet [197]. This difference developed despite the birds having been on the same diet continuously since 15 days of age, in fact for 85% of their lives. These results suggest that the quality of the rearing diet permanently affected the capacity of birds to assimilate lipophilic antioxidants from the diet. Effect of early nutrition in mammals is well recognised. In particular, retrospective studies investigating the effect of famine or season during pregnancy indicate that variation in early environmental exposure in utero leads to differences in DNA methylation of offspring [198]. This potentially may affect gene expression in the offspring via epigenetic mechanisms [175]. Our recent results demonstrated substantial effects of diet on the development of behavioural traits, and that these effects differ both between the sexes and over different developmental periods [199].
It seems likely that early programming associated with epigenetic mechanisms plays a key role in chicken growth and development and at time of chicken placement additional supplementation of fat-soluble vitamins, which are poorly assimilated from the diet, with water could be considered as an important solution for poultry industry.
Critical periods of the gut development: pig weaning
Piglets face physiological, environmental, and social stressful challenges when weaned from the sow. Indeed, the process of weaning is one of the most stressful events in the pig’s life that can contribute to intestinal and immune system dysfunctions resulting in reduced pig health, growth, and feed intake, particularly during the first week after weaning [200]. Indeed, the piglet digestive system faces a range of challenges associated with abrupt change from highly digestible and palatable liquid milk from its mother to a solid dry diet that is less digestible and palatable. This leads to reduced feed intake after weaning and the piglet becomes malnourished with reduced transient growth rate [200]. The weaning process is associated with marked structural and functional changes in the small intestine, including villous shedding, shortening and atrophy and crypt hyperplasia, which cause decreased digestive and absorptive capacity and contribute to post-weaning diarrhoea and poor performance [201].
Indeed, post-weaning anorexia and under-nutrition are major problems in post-weaning piglets. It seems likely that weaning is associated with changes in gene expression in the piglet gut. Recently, in an experiment conducted in China, changes in the genomic expression in the intestines of weaned pigs were studied by microarray analysis [202]. Four hundred forty-five genes showed altered expression after weaning treatment (286 upregulated and 159 downregulated). Most of the altered genes were responsible for regulation of various cellular processes associated with development and metabolism. The obtained results indicate that weaning induced cell cycle arrest, enhanced apoptosis, and inhibited epithelial cell proliferation, which might be important elements of molecular mechanisms of weaning stress. Wang et al. also showed decreased cell proliferation- and differentiation-related gene expression, in weaned pigs, and dietary additives such as plasma protein and arginine could enhance the crypt cell proliferation in weaned piglets [203,205]. A complexity of interactions between dietary components, the microbiota (or its metabolites), the enterocyte and the local immune system in piglets has been demonstrated [206,207]. There is a range of publications devoted to the development of management and feeding strategies to stimulate gut development and health in young piglets. The ultimate aim of these strategies is to prevent detrimental hanges in the gut and improve productivity of pigs around the time of weaning [208].
Similar to other species, including birds, antioxidant-prooxidant balance in the pig gut could be an important determinant of its functional ability in stress conditions of birth [209] and weaning [31]. It fact, piglet birth imposes a great oxidative stress and there are adaptive changes in the gut to overcome excessive free radical production. In particular, nuclear Nrf2 level gradually increased under birth oxidative stress in the ileum in early postnatal development. Indeed, with activation of Nrf2 signaling, expression of antioxidant genes, and the release of antioxidant enzymes, excessive ROS has been scavenged and birth oxidative balance gradually recovered [209]. Furthermore, it has been shown that weaning stress is also associated with oxidative stress in vivo, resulting in villus atrophy and reductions in the activities of digestive enzymes of weaned piglets [210].
In fact, weaning resulted in reductions in the villus height and width, and in the activity of digestive enzymes. Furthermore, activity of SOD decreased and the concentrations of MDA, NO, and H2O2 increased after weaning. It is interesting to note that the genes related to the antioxidant enzymes and digestive enzymes were down-regulated after weaning [210]. Similarly, the weaning stress caused a significant decrease in total antioxidant capacity in the piglet ileum, colon and cecum, GSH-Px activity in ileum and colon and increased lipid peroxidation in colon and cecum as well as increased H2O2 concentration in ileum, colon and cecum [211]. The same authors showed significant decreases in jejunum and colon antioxidant capacities, Lactobacillus and Bifidobacterium counts, and significant increases in levels of jejunum MDA, colon hydroxyl radicals, jejunum and colon H2O2, and Escherichia coli counts in weaned piglets [212]. The observed imbalance of the intestinal redox status and microbiota was significantly restored by the antioxidant blend, containing vitamins E, C and other antioxidant compounds. In newly weaned piglets, Zhu et al. observed the upregulation of p53, which regulates reactive oxygen species generation, and down-regulation of PPARγ coactivator-1α (PGC-1α), which plays an important protective role against oxidative stress by regulating the expression of mitochondrial antioxidants [210]. Furthermore, early weaning suppressed expression of p65 (at 3, 5, and 7 days post-weaning) and Nrf2 (at 5 and 7 d) in the piglet jejunum [213]. In general, aforementioned results clearly indicated that intestinal dysfunction after weaning is associated with an inhibition of the antioxidant system. The same authors [210] showed that antioxidant blend, containing vitamin E and C and other antioxidants, was able to decrease free radical-induced damage and suppress oxidative stress by modulating the expressions of tumor protein 53 and PGC- 1α genes. Taking into account those data it is possible to conclude that nutritional antioxidants (vitamins E, C, selenium, carnitine, etc.) could be key elements in overcoming detrimental changes in the gut of the newly weaned piglets. For example, Gessner observed a lower transactivation of NF-κB and a lower expression of various target genes of this transcription factor in the duodenal mucosa of pigs fed antioxidant-rich grape seed extract [214]. Positive effects of similar antioxidant-rich grape extract on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs has been recently confirmed [215]. Furthermore, early weaninginduced growth retardation appears to be attenuated through changes in plasma amino acid profiles and elevation of growth performance and intestinal antioxidant capacity in piglets following increased consumption of methionine as DL-2-hydroxy-4-methylthiobutyrate [216]. Another dietary antioxidant, N-Acetyl cysteine was also beneficial for preserving morphological integrity in weaned piglets via the regulation of cell apoptosis [217]. In contrast, feeding weaning pigs diets containing 10% thermally oxidized lipids appeared to impair their oxidative status [218]. Indeed, antioxidant-prooxidant redox balance in the piglet gut is an important determinant of the gut health.
The role of vitagenes in maintaining redox balance in the piglet gut has been described recently [219]. Furthermore, positive effects of an antioxidant composition, developed on the vita-gene concept and containing vitamins, minerals, amino acids, carnitine, betaine, etc. (PerforMax) supplied to weaning piglets with drinking water was described [220]. Taking into account our preliminary observations we hypothesised that antioxidant-rich composition (PerforMax) provided with drinking water could activate the vita-gene network resulting in increased antioxidant defences of the gut due to activation of Nrf2. At the same time it could decrease activity of NF-κB in the gut leading to reduced inflammation. Indeed, at time of weaning, when feed consumption is low a supplementation of the antioxidant compounds via drinking water could be an important approach to deal with aforementioned detrimental changes in the weaning piglet gut. Clearly, more research in this exciting area is needed.
Conclusions: Looking Ahead
The data presented above clearly indicated that weaning stress is the main reason for the loss of intestinal barrier function and gut inflammation/dysfunction in early-weaned pigs associated with promotion of free radical generation, decreased antioxidant defences and reduction in digestive enzyme activities [201,208,210]. Similarproblems with antioxidant defences of the gut of the newly-hatched chickens could be observed [31,54,74]. In this regard, usage of antioxidant blends is shown to have beneficial effects in decreasing such detrimental changes in the gut. However, it is necessary to takeinto account that antioxidant systems of the gut is quite complex and regulated at the levels of vitagenes operating in close relationship with a range of transcription factors, including Nrf2 and NF-κB. Therefore, the strategy of improving antioxidant defences of the gut of the newly weaned piglets or newly-hatched chicks should include several points:• Regulation of mitochondria function to decrease free radical production (carnitine, betaine, vitamin E, etc.)• Activation of vita-gene network, responsible for synthesis of a range protective compounds, including Hsps, elements of the thioredoxin system, sirtuins, etc. (carnitine, betaine, vitamins A and E, etc.)
• Activation of Nrf2, responsible for synthesis of antioxidant and detoxification enzymes
• Suppression of NF-κB, responsible for synthesis of proinflammatoryz cytokine
• Provision of vitamin E and elements of its biological recycling in the cell (vitamin C, selenium, vitamin B2, etc.), since vitamin E is major membrane antioxidant, which cannot be replaced by other antioxidants
• Provision of minerals necessary for additional synthesis of SOD and GSH-Px (Zn, Mn, Se)
There is a great body of evidence indicating that during pig weaning and in the first days of the chicken postnatal development many aforementioned elements are compromised (see sections above). In addition, the plasma levels of betaine, creatine, L-arginine and acetyl-carnitine were lower on day 4 post-weaning than on day 0 [221] and antioxidant action of carnitine and betaine received a substantial attention for the last decade [222,228]. Indeed, a design and effective usage of the complex antioxidant composition to overcome oxidative stress related to weaning in piglets or chick placement is an important task for nutritionists.
References
- Diplock AT (1994) Antioxidants and disease prevention. Mol Aspects Med 15: 293-376.
- Staprans I, Hardman DA, Pan XM, Feingold KR (1999) Effect of oxidized lipids in the diet on oxidized lipid levels in postprandial serum chylomicrons of diabetic patients. Diabetes Care 22: 300-306.
- Staprans I, Rapp JH, Pan XM, Kim KY, Feingold KR (1994) Oxidized lipids in the diet are a source of oxidized lipid in chylomicrons of human serum. Arterioscler Thromb 14: 1900-1905.
- Hayam I, Cogan U, Mokady S (1997) Enhanced peroxidation of proteins of the erythrocyte membrane and of muscle tissue by dietary oxidized oil. Biosci Biotechnol Biochem 61: 1011-1012.
- Lin CF, Asghar A, Gray JI, Buckley DJ, Booren AM, et al. (1989) Effects of oxidized dietary oil and antioxidant supplementation on broiler growth and meat stability. Br Poult Sci 30: 855-864.
- Calabotta DF, Shermer WD (1985) Controlling feed oxidation can be rewarding. Feedstuffs 57: 24-33.
- Sheehy PJ, Morrissey PA, Flynn A (1994) Consumption of thermally-oxidized sunflower oil by chicks reduces alpha-tocopherol status and increases susceptibility of tissues to lipid oxidation. Br J Nutr 71: 53-65.
- Yang CM, Kendall CW, Stamp D, Medline A, Archer MC, et al. (1998) Thermally oxidized dietary fat and colon carcinogenesis in rodents. Nutr Cancer 30: 69-73.
- Andia AM, Street JC (1975) Dietary induction of hepatic microsomal enzymes by thermally oxidized fats. J Agric Food Chem 23: 173-177.
- Cutler MG, Schneider R (1973) Malformations produced in mice and rats by oxidized linoleate. Food Cosmet Toxicol 11: 935-939.
- Koshio S, Ackmman RG, Lall SP (1994) Effects of oxidized herring and canola oil diets on growth survival and flavour of Atlantic salmon, Salmon salar. J Agric Food Chem 42: 1164-1169.
- Shibata K, Onodera M, Ashida H, Kanadawa K (1992) Effects of peroxidation products of linoleic acid on tryptophan-nicotinamide metabolism in rats. Biosci Biotechnol Biochem 56: 1270-1274.
- Liu P, Kerr BJ, Weber TE, Chen C, Johnston LJ, et al. (2014) Influence of thermally oxidized vegetable oils and animal fats on intestinal barrier function and immune variables in young pigs. J Anim Sci 92: 2971-2979.
- Liu P, Chen C, Kerr BJ, Weber TE, Johnston LJ, et al. (2014) Influence of thermally oxidized vegetable oils and animal fats on growth performance, liver gene expression, and liver and serum cholesterol and triglycerides in young pigs. J Anim Sci 92: 2960-2970.
- Luci S, König B, Giemsa B, Huber S, Hause G, et al. (2007) Feeding of a deep-fried fat causes PPARalpha activation in the liver of pigs as a non-proliferating species. Br J Nutr 97: 872-882.
- Dibner JJ, Atwell CA, Kitchell ML, Shermer WD, Ivey FJ (1996) Feeding of oxidised fats to broilers and swine: effects on enterocyte turnover, hepatocyte proliferation and the gut associated lymphoid tissue. Anim Feed Sci Technol 62: 1-13.
- Great Britain. Panel on Dietary Reference Values (1991) Dietary reference values for food energy and nutrients for the United Kingdom: Report of the Panel on Dietary Reference Values of the Committee on Medical Aspects of Food Policy. H.M. Stationery Office, London.
- Davies MJ, Slater TF (1987) Studies on the metal-ion and lipoxygenase-catalysed breakdown of hydroperoxides using electron-spin-resonance spectroscopy. Biochem J 245: 167-173.
- Braughler JM, Duncan LA, Chase RL (1986) The involvement of iron in lipid peroxidation. Importance of ferric to ferrous ratios in initiation. J Biol Chem 261: 10282-10289.
- Minotti G, Aust SD (1987) The role of iron in the initiation of lipid peroxidation. Chem Phys Lipids 44: 191-208.
- Hollan S, Johansen KS (1993) Adequate iron stores and the 'Nil nocere' principle. Haematologia (Budap) 25: 69-84.
- Knutson MD, Walter PB, Ames BN, Viteri FE (2000) Both iron deficiency and daily iron supplements increase lipid peroxidation in rats. J Nutr 130: 621-628.
- Reifen R, Matas Z, Zeidel L, Berkovitch Z, Bujanover Y (2000) Iron supplementation may aggravate inflammatory status of colitis in a rat model. Dig Dis Sci 45: 394-397.
- Kadiiska MB, Burkitt MJ, Xiang QH, Mason RP (1995) Iron supplementation generates hydroxyl radical in vivo. An ESR spin-trapping investigation. J Clin Invest 96: 1653-1657.
- Blakeborough MH, Owen RW, Bilton RF (1989) Free radical generating mechanisms in the colon: their role in the induction and promotion of colorectal cancer? Free Radic Res Commun 6: 359-367
- Buishi IE (2004) Epidemiology of canine echinococcosis in northwest Libya, northwest Kenya and mid-Wales UK: Ph.D. Thesis in Parasitology, Bioscience Research Institute, School of Environment and life Science University of Salford.
- Chow CK, Hong CB (2002) Dietary vitamin E and selenium and toxicity of nitrite and nitrate. Toxicology 180: 195-207.
- Benjamin N, O'Driscoll F, Dougall H, Duncan C, Smith L, et al. (1994) Stomach NO synthesis. Nature 368: 502.
- Zweier JL, Samouilov A, Kuppusamy P (1999) Non-enzymatic nitric oxide synthesis in biological systems. Biochim Biophys Acta 1411: 250-262.
- Van Dyke K (1997) The possible role of peroxynitrite in Alzheimer's disease: a simple hypothesis that could be tested more thoroughly. Med Hypotheses 48: 375-389
- Surai PF (2006) Selenium in nutrition and health. Nottingham University Press, Nottingham, United Kingdom.
- Khandelwal S, Shukla LJ, Shanker R (2002) Modulation of acute cadmium toxicity by Emblica officinalis fruit in rat. Indian J Exp Biol 40: 564-570.
- Wang X, Yin X, Bai X (2002) Combined effect of lead and cadmium on lipid peroxidation in renal tubular epithelial cells of rats. Wei Sheng Yan Jiu 31: 232-234.
- Dew HR (1928) Hydatid disease its pathology, diagnosis and treatment. Sydeney. The Australian Medical Publishing Company Limited.
- Chen L, Yang X, Jiao H, Zhao B (2002) Tea catechins protect against lead-induced cytotoxicity, lipid peroxidation, and membrane fluidity in HepG2 cells. Toxicol Sci 69: 149-156
- Ercal N, Neal R, Treeratphan P, Lutz PM, Hammond TC, et al. (2000) A role for oxidative stress in suppressing serum immunoglobulin levels in lead-exposed Fisher 344 rats. Arch Environ Contam Toxicol 39: 251-256.
- Upasani CD, Khera A, Balaraman R (2001) Effect of lead with vitamin E, C, or Spirulina on malondialdehyde, conjugated dienes and hydroperoxides in rats. Indian J Exp Biol 39: 70-74.
- Mahboob M, Shireen KF, Atkinson A, Khan AT (2001) Lipid peroxidation and antioxidant enzyme activity in different organs of mice exposed to low level of mercury. J Environ Sci Health B 36: 687-697.
- Jensen E, Egan S, Canady R, Bolger P (2001) Dietary exposures to persistent organic pollutants. Toxicol Ind Health 17: 157-162.
- Furusawa N, Morita Y (2000) Polluting profiles of dieldrin and DDTs in laying hens of Osaka, Japan. J Vet Med B Infect Dis Vet Public Health 47: 511-515.
- Lovell RA, McChesney DG, Price WD (1996) Organohalogen and organophosphorus pesticides in mixed feed rations: findings from FDA's domestic surveillance during fiscal years 1989-1994. J AOAC Int 79: 544-548.
- Nag SK, Raikwar MK (2011) Persistent organochlorine pesticide residues in animal feed. Environ Monit Assess 174: 327-335.
- Latchoumycandane C, Mathur PP (2002) Induction of oxidative stress in the rat testis after short-term exposure to the organochlorine pesticide methoxychlor. Arch Toxicol 76: 692-698.
- Latchoumycandane C, Mathur PP (2002) Induction of oxidative stress in the rat testis after short-term exposure to the organochlorine pesticide methoxychlor. Arch Toxicol 76: 692-698.
- Sahoo A, Samanta L, Chainy GB (2000) Mediation of oxidative stress in HCH-induced neurotoxicity in rat. Arch Environ Contam Toxicol 39: 7-12.
- Fink-Gremmels J (1999) Mycotoxins: their implications for human and animal health. Vet Q 21: 115-120.
- Surai P, Mezes M, Fotina TI, Denev SD (2010) Mycotoxins in human diet: a hidden danger. In: Modern Dietary Fat Intakes in Disease Promotion (Fabien De Meester, Sherma Zibadi, Ronald Ross Watson, Eds). Humana Press, Springer New York London, pp. 275-303.
- Sherazi ST, Shar ZH, Sumbal GA, Tan ET, Bhanger MI, et al. (2015) Occurrence of ochratoxin A in poultry feeds and feed ingredients from Pakistan. Mycotoxin Res 31: 1-7.
- Greco MV, Franchi ML, Rico Golba SL, Pardo AG, Pose GN (2014) Mycotoxins and mycotoxigenic fungi in poultry feed for food-producing animals. ScientificWorldJournal 2014: 968215 .
- Li X, Zhao L, Fan Y, Jia Y, Sun L, et al. (2014) Occurrence of mycotoxins in feed ingredients and complete feeds obtained from the Beijing region of China. J Anim Sci Biotechnol 5: 37.
- Wang Y, Liu S, Zheng H, He C, Zhang H (2013) T-2 toxin, zearalenone and fumonisin B₁ in feedstuffs from China. Food Addit Contam Part B Surveill 6: 116-122.
- Streit E, Naehrer K, Rodrigues I, Schatzmayr G (2013) Mycotoxin occurrence in feed and feed raw materials worldwide: long-term analysis with special focus on Europe and Asia. J Sci Food Agric 93: 2892-2899.
- Maresca M, Mahfoud R, Pfohl-Leszkowicz A, Fantini J (2001) The mycotoxin ochratoxin A alters intestinal barrier and absorption functions but has no effect on chloride secretion. Toxicol Appl Pharmacol 176: 54-63.
- Maresca M, Mahfoud R, Garmy N, Fantini J (2002) The mycotoxin deoxynivalenol affects nutrient absorption in human intestinal epithelial cells. J Nutr 132: 2723-2731.
- Surai PF (2002) Natural antioxidants in avian nutrition and reproduction. Nottingham University Press, Nottingham, UK.
- Surai PF, Dvorska JE (2005) Effects of mycotoxins on antioxidant status and immunity. In: The Mycotoxin Blue Book (Diaz DE, Ed.), Nottingham University Press, Nottingham, pp. 93-137.
- Surai PF, Mezes M, Melnichuk S, Fotina T (2008) Mycotoxins and animal health: from oxidative stress to gene expression. Krmiva 50: 35-43.
- Awad W, Ghareeb K, Böhm J, Zentek J (2013) The toxicological impacts of the Fusarium mycotoxin, deoxynivalenol, in poultry flocks with special reference to immunotoxicity. Toxins (Basel) 5: 912-925.
- Maresca M (2013) From the gut to the brain: journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins (Basel) 5: 784-820.
- Grenier B, Applegate TJ (2013) Modulation of intestinal functions following mycotoxin ingestion: meta-analysis of published experiments in animals. Toxins (Basel) 5: 396-430.
- Smith LE, Stoltzfus RJ, Prendergast A (2012) Food chain mycotoxin exposure, gut health, and impaired growth: a conceptual framework. Adv Nutr 3: 526-531.
- Pestka JJ (2010) Deoxynivalenol: mechanisms of action, human exposure, and toxicological relevance. Arch Toxicol 84: 663-679.
- Halliwell B, Zhao K, Whiteman M (2000) The gastrointestinal tract: a major site of antioxidant action? Free Radic Res 33: 819-830.
- Maaser C, Kagnoff MF (2002) Role of the intestinal epithelium in orchestrating innate and adaptive mucosal immunity. Z Gastroenterol 40: 525-529.
- Kiyono H, Kweon MN, Hiroi T, Takahashi I (2001) The mucosal immune system: from specialized immune defense to inflammation and allergy. Acta Odontol Scand 59: 145-153.
- Laroux FS, Pavlick KP, Wolf RE, Grisham MB (2001) Dysregulation of intestinal mucosal immunity: implications in inflammatory bowel disease. News Physiol Sci 16: 272-277.
- Nagura H, Ohtani H, Sasano H, Matsumoto T (2001) The immuno-inflammatory mechanism for tissue injury in inflammatory bowel disease and Helicobacter pylori-infected chronic active gastritis. Roles of the mucosal immune system. Digestion 63 Suppl 1: 12-21.
- Cunningham-Rundles S (2001) Nutrition and the mucosal immune system. Curr Opin Gastroenterol 17: 171-176.
- Sanderson IR (1999) The physicochemical environment of the neonatal intestine. Am J Clin Nutr 69: 1028S-1034S.
- Kanner J, Lapidot T (2001) The stomach as a bioreactor: dietary lipid peroxidation in the gastric fluid and the effects of plant-derived antioxidants. Free Radic Biol Med 31: 1388-1395.
- Surai PF (1999) Vitamin E in avian reproduction. Poult Avi Biol Rev 10: 1-60
- Surai PF, Fisinin VI (2012) Modern methods of fighting stresses in poultry production: from antioxidants to vitagenes. Agri Biol (Selskokhozaistvennaya Biologia, Russia), 4: 3-13
- Fotina AA, Fisinin VI, Surai PF (2013) Recent developments in usage of natural antioxidants to improve chicken meat production and quality. Bulgarian J Agric Sci 19: 889-896.
- Surai PF (2014) Polyphenol compounds in the chicken/animal diet: from the past to the future. J Anim Physiol Anim Nutr (Berl) 98: 19-31.
- Surai PF (2014) Antioxidant systems of the body: from vitamin E to polyphenols and beyond. Proceedings of the 35th Western Nutrition Conference, Edmonton, Canada, pp. 265-277.
- McLean JA, Karadas F, Surai PF, McDevitt RM, Speake BK (2005) Lipid-soluble and water-soluble antioxidant activities of the avian intestinal mucosa at different sites along the intestinal tract. Comp Biochem Physiol B Biochem Mol Biol 141: 366-372.
- Mocchegiani E, Costarelli L, Giacconi R, Malavolta M, Basso A, et al. (2014) Vitamin E-gene interactions in aging and inflammatory age-related diseases: implications for treatment. A systematic review. Ageing Res Rev 14: 81-101.
- Overvad K, Diamant B, Holm L, Holmer G, Mortensen SA, et al. (1999) Coenzyme Q10 in health and disease. Eur J Clin Nutr 53: 764-770.
- Ernster L, Dallner G (1995) Biochemical, physiological and medical aspects of ubiquinone function. Biochim Biophys Acta 1271: 195-204.
- Navarro F, Navas P, Burgess JR, Bello RI, De Cabo R, et al. (1998) Vitamin E and selenium deficiency induces expression of the ubiquinone-dependent antioxidant system at the plasma membrane. FASEB J 12: 1665-1673.
- Grudzinski IP, Frankiewicz-Jozko A (2001) Effects of oral coenzyme Q10 supplementation on sodium nitrite-induced lipid peroxidation in rats. Rocz Panstw Zakl Hig 52: 213-218.
- Matsusaka C, Marubayashi S, Dohi K, Kawasaki T (1992) The protective effect of administered CoQ10 against small intestinal damage caused by ischemia reperfusion. Transplant Proc 24: 1090-1091.
- Novoselova EG, Kolomiitseva IK, Obol'nikova EA, Samokhvalov GI, Kuzin AM (1985) Effect of ubiquinone on phospholipid metabolism in radiation injury. Biull Eksp Biol Med 99: 440-442.
- Buffinton GD, Doe WF (1995) Depleted mucosal antioxidant defences in inflammatory bowel disease. Free Radic Biol Med 19: 911-918.
- Maoka T (2009) Recent progress in structural studies of carotenoids in animals and plants. Arch Biochem Biophys 483: 191-195.
- Surai PF (2012) The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 1. Worlds Poult Sci J 68: 465-476.
- Surai PF (2012) The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 2 68: 717- 726 .
- Serpeloni JM, Cólus IM, de Oliveira FS, Aissa AF, Mercadante AZ, et al. (2014) Diet carotenoid lutein modulates the expression of genes related to oxygen transporters and decreases DNA damage and oxidative stress in mice. Food Chem Toxicol 70: 205-213.
- 88. Hathcock JN (1997) Vitamins and minerals: efficiency and safety. Am J Clin Nutr 66: 427-437.
- Loguercio C, Di Pierro M (1999) The role of glutathione in the gastrointestinal tract: a review. Ital J Gastroenterol Hepatol 31: 401-407.
- Hansen JM, Harris C (2014) Glutathione during embryonic development. Biochim Biophys Acta [Epub ahead of print].
- Aquilano K, Baldelli S, Ciriolo MR (2014) Glutathione: new roles in redox signaling for an old antioxidant. Front Pharmacol 5: 196.
- Ribas V, García-Ruiz C, Fernández-Checa JC (2014) Glutathione and mitochondria. Front Pharmacol 5: 151.
- Griffiths HR, Dias IH, Willetts RS, Devitt A (2014) Redox regulation of protein damage in plasma. Redox Biol 2: 430-435.
- Circu ML, Aw TY (2011) Redox biology of the intestine. Free Radic Res 45: 1245-1266.
- Aw TY (2005) Intestinal glutathione: determinant of mucosal peroxide transport, metabolism, and oxidative susceptibility. Toxicol Appl Pharmacol 204: 320-328.
- 96. Jefferies H, Bot J, Coster J, Khalil A, Hall JC, et al. (2003) The role of glutathione in intestinal dysfunction. J Invest Surg 16: 315-323..
- Aw TY (1998) Determinants of intestinal detoxication of lipid hydroperoxides. Free Radic Res 28: 637-646.
- Pietta PG (2000) Flavonoids as antioxidants. J Nat Prod 63: 1035-1042.
- Dolara P, Luceri C, De Filippo C, Femia A P, Giovannelli L, et al. (2005) Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat Res 591: 237-246.
- Giovannelli L, Testa G, De Filippo C, Cheynier V, Clifford MN, et al. (2000) Effect of complex polyphenols and tannins from red wine on DNA oxidative damage of rat colon mucosa in vivo. Eur J Nut 39: 207-212.
- Gorelik S, Lapidot T, Shaham I, Granit R, Ligumsky M, et al. (2005) Lipid peroxidation and coupled vitamin oxidation in simulated and human gastric fluid inhibited by dietary polyphenols: health implications. J Agric Food Chem 53: 3397-3402.
- Circu ML, Aw TY (2012) Intestinal redox biology and oxidative stress. Semin Cell Dev Biol 23: 729-737.
- Martínez JA, Etxeberría U, Galar A, Milagro FI (2013) Role of dietary polyphenols and inflammatory processes on disease progression mediated by the gut microbiota. Rejuvenation Res 16: 435-437.
- Biasi F, Deiana M, Guina T, Gamba P, Leonarduzzi G, et al. (2014) Wine consumption and intestinal redox homeostasis. Redox Biol 2: 795-802..
- Biasi F, Astegiano M, Maina M, Leonarduzzi G, Poli G (2011) Polyphenol supplementation as a complementary medicinal approach to treating inflammatory bowel disease. Curr Med Chem 18: 4851-4865.
- Lu T, Harper AF, Dibner JJ, Scheffler JM, Corl BA, et al. (2014) Supplementing antioxidants to pigs fed diets high in oxidants: II. Effects on carcass characteristics, meat quality, and fatty acid profile. J Anim Sci 92: 5464-5475.
- Lu T, Harper AF, Zhao J, Estienne MJ, Dalloul RA (2014) Supplementing antioxidants to pigs fed diets high in oxidants: I. Effects on growth performance, liver function, and oxidative status. J Anim Sci 92: 5455-5463.
- Lu T, Harper AF, Zhao J, Dalloul RA (2014) Effects of a dietary antioxidant blend and vitamin E on growth performance, oxidative status, and meat quality in broiler chickens fed a diet high in oxidants. Poult Sci 93: 1649-1657.
- Lu T, Harper AF, Zhao J, Corl BA, LeRoith T, et al. (2014) Effects of a dietary antioxidant blend and vitamin E on fatty acid profile, liver function, and inflammatory response in broiler chickens fed a diet high in oxidants. Poult Sci 93: 1658-1666.
- Chu FF, Doroshow JH, Esworthy RS (1993) Expression, characterization, and tissue distribution of a new cellular selenium-dependent glutathione peroxidase, GSHPx-GI. J Biol Chem 268: 2571-2576.
- Esworthy RS, Swiderek KM, Ho YS, Chu FF (1998) Selenium-dependent glutathione peroxidase-GI is a major glutathione peroxidase activity in the mucosal epithelium of rodent intestine. Biochim Biophys Acta 1381: 213-226.
- Brigelius-Flohe R (1999) Tissue-specific functions of individual glutathione peroxidases. Free Radic Biol Med 27: 951-965.
- Brigelius-Flohé R, Maiorino M (2013) Glutathione peroxidases. Biochim Biophys Acta 1830: 3289-3303.
- Mork H, Lex B, Scheurlen M, Dreher I, Schutze N, et al. (1998) Expression pattern of gastrointestinal selenoproteins--targets for selenium supplementation. Nutr Cancer 32: 64-70.
- LeGrand TS, Aw TY (1998) Chronic hypoxia alters glucose utilization during GSH-dependent detoxication in rat small intestine. Am J Physiol 274: G376-G384.
- Rattan SI (1998) The nature of gerontogenes and vitagenes. Antiaging effects of repeated heat shock on human fibroblasts. Ann N Y Acad Sci 854: 54-60.
- Calabrese V, Boyd-Kimball D, Scapagnini G, Butterfield DA (2004) Nitric oxide and cellular stress response in brain aging and neurodegenerative disorders: the role of vitagenes. In Vivo 18: 245-267.
- Calabrese V, Guagliano E, Sapienza M, Panebianco M, Calafato S, et al. (2007) Redox regulation of cellular stress response in aging and neurodegenerative disorders: role of vitagenes. Neurochem Res 32: 757-773.
- Calabrese V, Scapagnini G, Davinelli S, Koverech G, Koverech A, et al. (2014) Sex hormonal regulation and hormesis in aging and longevity: role of vitagenes. J Cell Commun Signal 8: 369-384.
- Cornelius C, Graziano A, Calabrese EJ, Calabrese V (2013) Hormesis and vitagenes in aging and longevity: mitochondrial control and hormonal regulation. Horm Mol Biol Clin Investig 16: 73-89.
- Cornelius C, Trovato Salinaro A, Scuto M, Fronte V, Cambria MT, et al. (2013) Cellular stress response, sirtuins and UCP proteins in Alzheimer disease: role of vitagenes. Immun Ageing 10: 41.
- Cornelius C, Perrotta R, Graziano A, Calabrese EJ, Calabrese V (2013) Stress responses, vitagenes and hormesis as critical determinants in aging and longevity: Mitochondria as a "chi". Immun Ageing 10: 15.
- Surai PF, Fisinin VI (2012) The modern antistress technologies in poultry: From antioxidants to vitagenes. Sel'skokhozyaistvennaya Biologiya (Agric Biol, Moscow) 4: 3-13.
- Fisinin VI, Surai PF (2011) Effective protection from stresses in poultry production: from vitamins to vitagenes. Part 1. Ptitza I Ptitzeproducti (Poult Poult Prod, Moscow) 5: 23-26.
- Fisinin VI, Surai PF (2011) Effective protection from stresses in poultry production: from vitamins to vitagenes. Part 2. Ptitza I Ptitzeproducti (Poult Poult Prod, Moscow) 6: 10-13.
- Velichko OA, Shabaldin SV, Surai PF (2013) Practical aspects of vitagene concept usage in poultry production. Ptitza I Ptitzeproducti (Poult Poult Prod, Moscow) 4: 42-45
- Trovato Salinaro A, Cornelius C, Koverech G, Koverech A, Scuto M, et al. (2014) Cellular stress response, redox status, and vitagenes in glaucoma: a systemic oxidant disorder linked to Alzheimer's disease. Front Pharmacol 5: 129.
- Trovato Salinaro A, Cornelius C, Koverech G, Koverech A, Scuto M, et al. (2014) Cellular stress response, redox status, and vitagenes in glaucoma: a systemic oxidant disorder linked to Alzheimer's disease. Front Pharmacol 5: 129.
- Liu HY, Lundh T, Dicksved J, Lindberg JE (2012) Expression of heat shock protein 27 in gut tissue of growing pigs fed diets without and with inclusion of chicory fiber. J Anim Sci 90 Suppl 4: 25-27.
- David JC, Grongnet JF, Lalles JP (2002) Weaning affects the expression of heat shock proteins in different regions of the gastrointestinal tract of piglets. J Nutr 132: 2551-2561.
- Liu HY, Dicksved J, Lundh T, Lindberg JE (2014) Expression of heat shock proteins 27 and 72 correlates with specific commensal microbes in different regions of porcine gastrointestinal tract. Am J Physiol Gastrointest Liver Physiol 306: G1033-G1041.
- Kopecek P, Altmannova K, Weigl E (2001) Stress proteins: nomenclature, division and functions. Biomedical Papers of Medicine Faculty of University Palacky, Olomouc, Czech Republic 145: 39-47.
- Yenari MA (2002) Heat shock proteins and neuroprotection. Adv Exp Med Biol 513: 281-299.
- Wadhwa R, Taira K, Kaul SC (2002) An Hsp70 family chaperone, mortalin/ mthsp70/PBP74/Grp75: what, when, and where? Cell Stress Chaperones 7: 309-316.
- Tsukimi Y, Okabe S (2001) Recent advances in gastrointestinal pathophysiology: role of heat shock proteins in mucosal defense and ulcer healing. Biol Pharm Bull 24: 1-9.
- Graf PC, Jakob U (2002) Redox-regulated molecular chaperones. Cell Mol Life Sci 59: 1624-1631.
- Prabhu R, Balasubramanian KA (2002) Heat preconditioning attenuates oxygen free radical-mediated alterations in the intestinal brush border membrane induced by surgical manipulation. J Surg Res 107: 227-233.
- Malago JJ, Koninkx JF, van Dijk JE (2002) The heat shock response and cytoprotection of the intestinal epithelium. Cell Stress Chaperones 7: 191-199.
- Urayama S, Musch MW, Retsky J, Madonna MB, Straus D, et al. (1998) Dexamethasone protection of rat intestinal epithelial cells against oxidant injury is mediated by induction of heat shock protein 72. J Clin Invest 102: 1860-1865.
- Beck SC, Paidas CN, Mooney ML, Deitch EA, De Maio A (1995) Presence of the stress-inducible form of hsp-70 (hsp-72) in normal rat colon. Shock 3: 398-402.
- Rokutan K (1999) Molecular stress response in the stomach. Nippon Yakurigaku Zasshi 114: 265-272.
- Fujii H, Takahashi T, Nakahira K, Uehara K, Shimizu H, et al. (2003) Protective role of heme oxygenase-1 in the intestinal tissue injury in an experimental model of sepsis. Crit Care Med 31: 893-902.
- Godoy JR, Funke M, Ackermann W, Haunhorst P, Oesteritz S, et al. (2011) Redox atlas of the mouse. Immunohistochemical detection of glutaredoxin-, peroxiredoxin-, and thioredoxin-family proteins in various tissues of the laboratory mouse. Biochim Biophys Acta 1810: 2-92.
- Gasdaska JR, Gasdaska PY, Gallegos A, Powis G (1996) Human thioredoxin reductase gene localization to chromosomal position 12q23-q24.1 and mRNA distribution in human tissue. Genomics 37: 257-259.
- Kipp A, Banning A, van Schothorst EM, Méplan C, Schomburg L, et al. (2009) Four selenoproteins, protein biosynthesis, and Wnt signalling are particularly sensitive to limited selenium intake in mouse colon. Mol Nutr Food Res 53: 1561-1572.
- Kipp AP, Müller MF, Göken EM, Deubel S, Brigelius-Flohé R (2012) The selenoproteins GPx2, TrxR2 and TrxR3 are regulated by Wnt signalling in the intestinal epithelium. Biochim Biophys Acta 1820: 1588-1596.
- Sido B, Giese T, Autschbach F, Lasitschka F, Braunstein J, et al. (2005) Potential role of thioredoxin in immune responses in intestinal lamina propria T lymphocytes. Eur J Immunol 35: 408-417.
- Jaeger SU, Schroeder BO, Meyer-Hoffert U, Courth L, Fehr SN, et al. (2013) Cell-mediated reduction of human β-defensin 1: a major role for mucosal thioredoxin. Mucosal Immunol 6: 1179-1190.
- Takaishi S, Sawada M, Seno H, Kayahara T, Morita-Fujisawa Y, et al. (2003) Growth promoting effect of thioredoxin on intestinal epithelial cells. Dig Dis Sci 48: 379-385.
- Higashikubo A, Tanaka N, Noda N, Maeda I, Yagi K, et al. (1999) Increase in thioredoxin activity of intestinal epithelial cells mediated by oxidative stress. Biol Pharm Bull 22: 900-903.
- Decuypere E, Tona K, Bruggeman V, Bamelis F (2001) The day-old chick: a crucial hinge between breeders and broilers. Worlds Poult Sci J 57: 127-138.
- Noy Y, Sklan D (1999) Different types of early feeding and performance in chicks and poults. J Appl Poult Res 8: 16-24.
- Geyra A, Uni Z, Sklan D (2001) Enterocyte dynamics and mucosal development in the posthatch chick. Poult Sci 80: 776-782.
- Plnchasov Y, Noy Y (1993) Comparison of post-hatch holding time and subsequent early performance of broiler chicks and Turkey poults. Br Poultry Sci 34: 111-120.
- Hager JE, Beane WL (1983) Posthatch incubation time and early growth of broiler chickens. Poultry Sci 62: 247-254.
- Sklan D, Noy Y, Hoyzman A, Rozenboim I (2000) Decreasing weight loss in the hatchery by feeding chicks and poults in hatching trays. J Appl Poult Res 9: 142-148.
- Vieira S L, Moran ETJr (1999) Effects of delayed placement and used litter on broiler yields. J Appl Poult Res 8: 75-81.
- Surai PF, Ionov IA, Kuklenko TV, Kostjuk IA, MacPherson A, et al. (1998) Effect of supplementing the hen's diet with vitamin A on the accumulation of vitamins A and E, ascorbic acid and carotenoids in the egg yolk and in the embryonic liver. Br Poult Sci 39: 257-263.
- Surai PF, Noble RC, Speake BK (1996) Tissue-specific differences in antioxidant distribution and susceptibility to lipid peroxidation during development of the chick embryo. Biochim Biochys Acta 1304: 1-10.
- Noy Y, Gyra A, Sklan D (2001) The effect of early feeding on growth and small intestinal development in the posthatch poult. Poult Sci 80: 912-919.
- Bigot K, Mignon-Grasteau P, Picard M, Tesseraud S (2003) Effects of delayed feed intake on body, intestine and muscle development in neonate broilers. Poult Sci 85: 781-788.
- Geyra A, Uni Z, Sklan D (2001) The effect of fasting at different ages on growth and tissue dynamics in the small intestine of the young chick. Br J Nutr 86: 53-61.
- Karadas F, Surai PF, Sparks NH (2011) Changes in broiler chick tissue concentrations of lipid-soluble antioxidants immediately post-hatch. Comp Biochem Physiol A Mol Integr Physiol 160: 68-71.
- Surai PF (2000) Effect of selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. Br Poult Sci 41: 235-243.
- Surai PF, Ionov IA, Kuchmistova EF, Noble RC, Speake BK (1998) The relationship between the levels of a-tocopherol and carotenoids in the maternal feed, yolk and neonatal tissues: comparison between the chicken, turkey, duck and goose. J Sci Food Agri 76: 593-598.
- Surai PF, Noble R, Speake B (1999) Relationship between vitamin E content and susceptibility to lipid peroxidation in tissues of the newly hatched chick. Br Poult Sci 40: 406-410.
- Kebede N, Mekonnen H, Wossene A, Tilahun G (2009) Hydatidosis of slaughtered cattle in Wolaita Sodo Abattoir, southern Ethiopia. Trop Anim Health Prod 41: 629-633.
- Surai PF, Speake BK, Noble RC, Sparks NH (1999) Tissue-specific antioxidant profiles and susceptibility to lipid peroxidation of the newly hatched chick. Biol Trace Elem Res 68: 63-78.
- Soto-Salanova MF, Sell JL, Mallarino EG, Piquer FJ, Barker DL, et al. (1993) Research note: Vitamin E status of turkey poults as influenced by different dietary vitamin E sources, a bile salt, and an antioxidant. Poult Sci 72: 1184-1188.
- Marusich WL, DeRitter E, Ogrinz EF, Keating J, Mitrovic M, et al. (1975) Effect of supplemental vitamin E on control of rancidity in poultry meat. Poult Sci 54: 831-844.
- Sell JL, Amgel CR, Piquer FJ, Mallarino EG, Al-Batshan HA (1991) Development patterns of selected characteristics of the gastrointestinal tract of young turkeys. Poult Sci 70: 1200-1205.
- Freeman CP (1976) Digestion and absorption of fat. In: Digestion in the fowl. Boorman KN and Freeman CP, Eds. British Poultry Science Ltd., Edinburgh pp. 117-142
- Sklan D, Bartov I, Hurwitz S (1982) Tocopherol absorption and metabolism in the chick and turkey. J Nutr 112: 1394-1400.
- Noy Y, Sklan D (1998) Yolk utilization in hatching birds. Proceedings of 10th European Poultry Conference, Jerusalem, Israel, pp. 435-438
- Amarasekera M, Prescott SL, Palmer DJ (2013) Nutrition in early life, immune-programming and allergies: the role of epigenetics. Asian Pac J Allergy Immunol 31:175-182.
- Sell JL (1996) Recent developments in vitamin E nutrition of turkeys. Anim Feed Sci Technol 60: 229-240.
- Applegate TJ, Sell JL (1996) Effect of dietary linoleic to linolenic acid ratio and vitamin E supplementation on vitamin E status of poults. Poult Sci 75: 881-890.
- Soto-Salanova MF, Sell JL (1995) Influence of supplemental dietary fat on changes in vitamin E concentration in livers of poults. Poult Sci 74: 201-204.
- Soto-Salanova MF, Sell JL (1996) Efficacy of dietary and injected vitamin E for poults. Poultry Sci 75: 1393-1403.
- Soto-Salanova MF (1998) Vitamin E in young turkeys: A reassessment of the requirement. Retrospective Theses and Dissertations, Digital Repository @ Iowa State University.
- Mezes M (1994) Effect of vitamin E treatment on early postnatal changes of vitamin E status of chicken. Acta Vet Hung 42: 477-480.
- Fisinin VI, Surai PF (2012) First days of chicken life: from a protection against stresses to an effective adaptation. Russian Poul Sci (Ptitsevodstvo, Russia) 2: 11-15
- 183. Fotina AA, Fotina TI, Surai PF (2011) Effect of antistress composition Feed-Food Magic Antistress Mix on broiler chicks during growth period. Ann Sumy Nat Agrarian Uni 2: 158-162
- Fotina A, Fotina TI, Surai PF (2014) Effect of a water-soluble antistress composition on broiler chickens. Proceedings of the. XIVth European Poultry Conference, Stavanger, Norway p: 555
- Velichko O, Surai PF (2014) Effect of an antistress composition supplied with water on chick growth and development. Proceedings of the XIVth European Poultry Conference, Stavanger, Norway p: 551
- Surai PF, Fotina AA, Fotina TI (2012) Effect of Feed Food Magic Antistress Mix on natural disease resistance of ducklings. Ann Sumy Nat Agrarian Uni (Ukraine) 7: 58-61
- Fisinin VI, Surai PF (2013) Immunity in modern animal and poultry production: from theory to practical aspects of immunomodulation. Russian Poult Sci (Ptitsevodstvo, Russia) 5: 4-10
- Fisinin VI, Surai PF (2013) Gut immunity in birds: facts and reflections. Agric Biol (Selskokhozaistvennaya Biologia, Russia) 4: 3-25.
- Fisinin VI, Surai PF (2012) Properties and toxicity of DON. Mycotoxins and antioxidants: Uncompromising fighting. Part 1. Anim Prod Russia (Zhivotnovodstvo Rossii) 5: 11-14
- Fisinin VI, Surai PF (2012) Properties and toxicity of DON. Mycotoxins and antioxidants: Uncompromising fighting. Part 2. Ani Prod Russia (Zhivotnovodstvo Rossii, Russia) 6: 3-5
- Fisinin VI, Surai PF (2012) Mycotoxins and antioxidants: Uncompromising fighting. Ochratoxin A. Part 1. Compounded Feed (Kombikorma, Russia) 3: 55-60
- Fisinin VI, Surai PF (2012) Mycotoxins and antioxidants: Uncompromising fighting. Ochratoxin A. Part 2. Compounded Feed (Kombikorma, Russia) 5: 59-60
- Fisinin VI, Surai PF (2012) Mycotoxins and antioxidants: Uncompromising fighting. T-toxin - metabolism and toxicity. Part 1. Poult Poult Prod (Ptiza i Ptizeproducti, Russia) 3: 38-41
- Fisinin VI, Surai PF (2012). Mycotoxins and antioxidants: Uncompromising fighting. T-toxin - mechanisms of toxicity and protection. Part 2. Poult Poult Prod (Ptiza i Ptizeproducti, Russia) 4: 36-39.
- 195. Surai PF, Fotina TI (2013). Physiological mechanisms of the heat stress development in poultry. Today’s Animal Production (Ukraine) 6: 54- 60.
- 196. Surai PF, Fotina TI (2013) Physiological mechanisms and practical approaches to decrease detrimental consequences of heat shock in pig production. Svinarstvo Ukraini (Pig Production in Ukraine) 25: 12-15.
- Blount JD, Metcalfe NB, Arnold KE, Surai PF, Devevey GL, et al. (2003) Neonatal nutrition, adult antioxidant defences and sexual attractiveness in the zebra finch. Proc Biol Sci 270: 1691-1696.
- Dominguez-Salas P, Cox SE, Prentice AM, Hennig BJ, Moore SE (2012) Maternal nutritional status, C(1) metabolism and offspring DNA methylation: a review of current evidence in human subjects. Proc Nutr Soc 71:154-165.
- Noguera JC, Metcalfe NB, Surai PF, Monaghan P (2015) Are you what you eat? Micronutritional deficiencies during development influence adult personality-related traits. Anim Behav 101: 129-140.
- Campbell JM, Crenshaw JD, Polo J (2013) The biological stress of early weaned piglets. J Anim Sci Biotechnol 4: 19.
- Pluske JR (2013) Feed- and feed additives-related aspects of gut health and development in weanling pigs. J Anim Sci Biotechnol 4: 1.
- Zhu LH, Xu JX, Zhu SW, Cai X, Yang SF, et al. (2014) Gene expression profiling analysis reveals weaning-induced cell cycle arrest and apoptosis in the small intestine of pigs. J Anim Sci 92: 996-1006.
- Wang J, Chen L, Li P, Li X, Zhou H, et al. (2008) Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J Nutr 138:1025-1032.
- Jiang R, Chang X, Stoll B, Fan MZ, Arthington J, et al. (2000) Dietary plasma protein reduces small intestinal growth and lamina propria cell density in early weaned pigs. J Nutr 130: 21-26.
- Liu Y, Huang J, Hou Y, Zhu H, Zhao S, et al. (2008) Dietary arginine supplementation alleviates intestinal mucosal disruption induced by Escherichia coli lipopolysaccharide in weaned pigs. Br J Nutr 100: 552-560.
- Lallès JP, Bosi P, Smidt H, Stokes CR (2007) Nutritional management of gut health in pigs around weaning. Proc Nutr Soc 66: 260-268.
- Lallès JP, Bosi P, Smidt H, Stokes CR (2007) Weaning - A challenge to gut physiologists. Livestock Sci 108: 82-93.
- De Lange CFM, Pluske J, Gong J, Nyachoti CM (2010) Strategic use of feed ingredients and feed additives to stimulate gut health and development in young pigs. Livestock Sci 134: 124-134.
- Yin J, Wu MM, Xiao H, Ren WK, Duan JL, et al. (2014) Development of an antioxidant system after early weaning in piglets. J Anim Sci 92: 612-619.
- Zhu LH, Zhao KL, Chen XL, Xu JX (2012) Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J Anim Sci 90: 2581-2589.
- Xu CC, Yang SF, Zhu LH, Cai X, Sheng YS, et al. (2014) Regulation of N-acetyl cysteine on gut redox status and major microbiota in weaned piglets. J Anim Sci 92: 1504-1511.
- Xu J, Xu C, Chen X, Cai X, Yang S, et al. (2014) Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets. Nutrition 30: 584-589.
- Yin J, Ren W, Liu G, Duan J, Yang G, et al. (2013) Birth oxidative stress and the development of an antioxidant system in newborn piglets. Free Radic Res 47: 1027-1035.
- Gessner DK, Fiesel A, Most E, Dinges J, Wen G, et al. (2013) Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet Scand 55: 18.
- Fiesel A, Gessner DK, Most E, Eder K (2014) Effects of dietary polyphenol-rich plant products from grape or hop on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs. BMC Vet Res 10: 196.
- Li H, Wan H, Mercier Y, Zhang X, Wu C, et al. (2014) Changes in plasma amino acid profiles, growth performance and intestinal antioxidant capacity of piglets following increased consumption of methionine as its hydroxy analogue. Br J Nutr 112: 855-67.
- Zhu L, Cai X, Guo Q, Chen X, Zhu S, et al. (2013) Effect of N-acetyl cysteine on enterocyte apoptosis and intracellular signalling pathways' response to oxidative stress in weaned piglets. Br J Nutr 110: 1938-1947.
- Liu P, Kerr BJ, Weber TE, Chen C, Johnston LJ, et al. (2014) Influence of thermally oxidized vegetable oils and animal fats on intestinal barrier function and immune variables in young pigs. J Anim Sci 92: 2971-2979.
- Surai PF, Melnichuk SD (2012). Mechanisms of Anti-Stress Protection in Pig Farming. From Vitamins to Vitagenes, Svinovodstvo Ukrainy (Pig Production in Ukraine) 2: 10-15
- Gaponov IV, Fotina TI, Surai PF (2012) Physiological and technological stresses at piglet weaning. Protective effect of the antistress composition. Svinovodstvo Ukrainy (Pig Production in Ukraine) 13: 6-9.
- Sugiharto S, Hedemann MS, Lauridsen C (2014) Plasma metabolomic profiles and immune responses of piglets after weaning and challenge with E. coli. J Anim Sci Biotechnol 5: 17.
- Ganesan B, Anandan R, Lakshmanan PT (2011) Studies on the protective effects of betaine against oxidative damage during experimentally induced restraint stress in Wistar albino rats. Cell Stress Chaperon 16: 641-652.
- Valero T (2014) Mitochondrial biogenesis: pharmacological approaches. Curr Pharm Des 20: 5507-5509.
- Ribas GS, Vargas CR, Wajner M (2014) L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders. Gene 533: 469-476
- Jones LL, McDonald DA, Borum PR (2010) Acylcarnitines: role in brain. Prog Lipid Res 49: 61-75.
- Calabrese V, Cornelius C, Dinkova-Kostova AT, Calabrese EJ (2009) Vitagenes, cellular stress response, and acetylcarnitine: relevance to hormesis. Biofactors 35: 146-160
- Hagar H, Medany AE, Salam R, Medany GE, Nayal OA (2015) Betaine supplementation mitigates cisplatin-induced nephrotoxicity by abrogation of oxidative/nitrosative stress and suppression of inflammation and apoptosis in rats. Exp Toxicol Pathol 67: 133-141.
- Hagar H, Al Malki W (2014) Betaine supplementation protects against renal injury induced by cadmium intoxication in rats: role of oxidative stress and caspase-3. Environ Toxicol Pharmacol 37: 803-811.