
Journal of Pharmaceutics & Pharmacology
Download PDF
Research Article
Address for Correspondence: Francesca Fernandez-Enright, Illawarra Health and Medical Research Institute, School of Psychology, University of Wollongong, Northfields Avenue, Wollongong, 2522, NSW, Australia, Tel: (+61) 2 4221 3494; Fax: (+61) 2 4221 8130; E-mail: fernande@uow.edu.au
Citation: Andrews JL, Newell KA, Matosin N, Huang XF, Fernandez-Enright F. NMDA Receptor Antagonism by Phencyclidine Reduces NWASP and WAVE1 Protein Expression and Reduces Levels of Myelination Markers in the Prefrontal Cortex of Rats. J Pharmaceu Pharmacol. 2015;3(1): 8.
Copyright © 2015 Andrews JL, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Pharmaceutics & Pharmacology |ISSN: 2327-204X | Volume: 3, Issue: 1
Submission: 20 January 2015 | Accepted: 23 February 2015 | Published: : 28 February 2015
Animals
NMDA Receptor Antagonism by Phencyclidine Reduces NWASP and WAVE1 Protein Expression and Reduces Levels of Myelination Markers in the Prefrontal Cortex of Rats
Jessica L. Andrews1,2, Kelly A. Newell1,2, Natalie Matosin1,2, Xu-Feng Huang1,2, Francesca Fernandez-Enright1,2,3*
- 1Illawarra Health and Medical Research Institute, Faculty of Science, Medicine and Health, University of Wollongong, New South Wales 2522, Australia
- 2Schizophrenia Research Institute, 405 Liverpool Street, DarlinghurstNew South Wales, 2010, Australia
- 3School of Psychology, Faculty of Social Sciences, University ofWollongong, New South Wales 2522, Australia
Address for Correspondence: Francesca Fernandez-Enright, Illawarra Health and Medical Research Institute, School of Psychology, University of Wollongong, Northfields Avenue, Wollongong, 2522, NSW, Australia, Tel: (+61) 2 4221 3494; Fax: (+61) 2 4221 8130; E-mail: fernande@uow.edu.au
Citation: Andrews JL, Newell KA, Matosin N, Huang XF, Fernandez-Enright F. NMDA Receptor Antagonism by Phencyclidine Reduces NWASP and WAVE1 Protein Expression and Reduces Levels of Myelination Markers in the Prefrontal Cortex of Rats. J Pharmaceu Pharmacol. 2015;3(1): 8.
Copyright © 2015 Andrews JL, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Pharmaceutics & Pharmacology |ISSN: 2327-204X | Volume: 3, Issue: 1
Submission: 20 January 2015 | Accepted: 23 February 2015 | Published: : 28 February 2015
Abstract
N-methyl-D-aspartate (NMDA) receptor antagonism by perinatal phencyclidine (PCP) treatment leads to neuronal damage and causes long-term behavioural alterations in rodents. It is routinely used to model pathological processes in the brain that may be present in schizophrenia, such as alterations to dendritic development, and disruptions to myelin processes. Both of these processes occur during brain development and are highly implicated in the schizophrenia pathophysiology. Changes to the polymerization and reorganization of the actin cytoskeleton can have significant effects on the morphology and dynamics of the dendrites within the brain. Actin regulation is primarily regulated by neural Wiskott-Aldrich syndrome protein (NWASP), and WASP-family verprolin homology protein-1 (WAVE1). Here we have examined the role of actin related, cytoskeletal proteins NWASP and WAVE1 in a neurodevelopmental model of schizophrenia using PCP to determine if these signaling pathways are altered in the prefrontal cortex and hippocampus throughout different stages of neurodevelopment. Male Sprague Dawley rats were injected subcutaneously with PCP (10 mg/kg) or saline at postnatal days (PN) 7, 9 and 11. Rats (n=6/group) were sacrificed at PN 12, 5 weeks or 14 weeks. Relative expression levels of protein expression were examined in the prefrontal cortex and hippocampus of the treated rats. NWASP, WAVE1 and MBP were decreased (0.001≤p≤0.032) in the prefrontal cortex of PCP treated rats at PN12. At 5 weeks of age, NWASP was reduced in the prefrontal cortex (p=0.037) and WAVE1 was reduced in the hippocampus (p=0.006). At 14 weeks, there were no significant changes in any of the tested proteins (p>0.05). This is the first report of an alteration in NWASP and WAVE1 proteins in the rat brain, directly following NMDA receptor antagonism by PCP treatment in early development. These findings suggest that alterations in these important scaffolding related proteins may contribute to the development of deficits in myelination and cognitive performance in the brain.Keywords
Neural Wiskott-Aldrich syndrome protein; WASP-family verprolin homology protein-1; Neurodevelopmental animal model; Schizophrenia; Myelin markersIntroduction
Glutamate is the primary excitatory neurotransmitter in the brain and through its actions predominantly on NMDA receptors, plays a key role in neurodevelopmental processes such as neuronal migration, synaptogenesis and synaptic plasticity in both cortical and sub cortical regions [1]. Phencyclidine or 1-(1-phenylcyclohexyl) piperidine (also known as PCP) is a potent non-competitive NMDA receptor antagonist [2-4]. To a lesser extent, PCP also has a lower binding affinity for several other targets including dopaminergic D2, nicotinic and muscarinic cholinergic receptors, as well as μ, κ and σ classes of opiate receptors [5-8]. In healthy human subjects, administration of PCP induces hallucinations and delusions which are common symptoms of schizophrenia; furthermore the administration of PCP to schizophrenia patients exacerbates their positive symptoms [9], thus a number of pharmacological models of schizophrenia have been developed using the administration of PCP to rodents (for review see [9]). Since the effects of PCP treatment in rodents are translatable to humans and other higher order primates, the administration of PCP is now one of the best known pharmacological models of schizophrenia [1,11]. Administering perinatal PCP to postnatal rodents has both the construct validity and face validity to test and address the proposed neurodevelopmental origins of schizophrenia. The administration of PCP at postnatal day (PN) 7, 9 and 11, which broadly corresponds to the second trimester of pregnancy in humans [12], has consistently been shown to induce hyperlocomotion, reduce prepulse inhibition and impair social interactions in rodents in adulthood [13-15], as previously published by our research group. All these behaviors illustrate symptoms encountered in schizophrenia pathology [9]. Due to its effects on a multitude of important receptor targets known to be involved in the pathophysiology of schizophrenia, and the schizophrenia-like behaviors, and symptoms it induces in rodents and humans respectively, PCP administration in rodents is now one of the most commonly used models for studying the development of schizophrenia [3,16,17]. In the postnatal period, from about PN7 to 14, neurons are highly sensitive to the toxic effects of NMDA receptor antagonists [18]; however neurons are not the only brain cells that are affected by disruptions to NMDA receptor antagonists. Oligodendrocytes are also very sensitive to PCP toxicity during development [19], and since they play a significant role in axonal connectivity and conduction, disruption to these processes during the early stages of brain development could impact significantly on normal brain development in its later stages.In addition to oligodendrocytes playing a significant role in nerve conductivity and communication, postsynaptic dendritic spines are also largely responsible for information processing and storage in neurons, as well as synaptic plasticity [20], all of which critically underlie learning and memory processes. Within the dendrites, it is the configuration of the actin cytoskeleton that is largely responsible for the regulation of synaptic spine formation and morphology; thus changes to the polymerization and reorganization of the actin cytoskeleton can have a significant impact on the morphology and dynamics of dendritic spines, which could lead to cognitive dysfunction [21]
Polymerization, reorganization and branching of the actin cytoskeleton are primarily regulated by the actin-related proteins (Arp2/3) complex. The binding of the Arp2/3 complex to the side of an existing actin filament elicits the polymerization of actin, resulting in filament branching [22]. The Arp2/3 complex is primarily regulated by neural Wiskott-Aldrich syndrome protein (NWASP), and WASP-family verprolin homology protein-1 (WAVE1) [23]. Loss of hippocampal NWASP and Arp3 has been shown to significantly reduce both the number of dendritic spines and the number of excitatory synapses [24]. Furthermore, genetic knockout of WAVE1 leads to hippocampal and cortical dysfunction, through a reduction in spine density and alterations in synaptic transmission [25]. WAVE1 knockout mice demonstrate learning and memory deficits, and display reduced sensorimotor function [26].
Since neuronal changes in NWASP and WAVE1 expression have been shown to induce alterations in dendritic spine growth and synaptic transmission [24,25,27], which both underlie learning and memory processes; we examined the levels of expression of these proteins throughout development in the prefrontal cortex and hippocampus of rats following perinatal administration of the potent NMDA receptor antagonist PCP. Furthermore, we also examined levels of myelination related proteins myelin basic protein (MBP) and myelin oligodendrocyte glycoprotein (MOG), since NWASP and WAVE1 are also critically involved in oligodendrocyte differentiation and myelination [28,29] which are also implicated in cognition.
Materials and Methods
Timed pregnant Sprague Dawley rats were obtained at gestation day 14 from the Animal Resource Centre (Perth, WA, Australia). They were housed in environmentally controlled conditions at 22 °C in a 12:12 hour light dark cycle with free access to food and water. The day of birth was considered PN0. Pups were sexed on PN7 and were randomly assigned to PCP or saline groups. The female pups were kept in the litters, however only male rats were used in this study. Pups were weaned at PN24-28, and were housed in pairs according to treatment. This study was approved by the Animal Ethics Committee at the University of Wollongong (AE13/01), and was conducted according to the guidelines of the Australian code of Practice for the Care and Use of Animals for Scientific Purposes, conforming to the International Guiding Principles for Biomedical Research Involving Animals. All efforts were made to minimize numbers of animals used and their suffering.
male Sprague Dawley rat pups were given a subcutaneous injection on PN7, 9 and 11; of PCP (10 mg/kg/day; Sigma, Castle Hill, NSW, Australia) or saline (0.9% NaCl at a volume of 1 ml/kg), as our group and others have previously shown that this treatment regime produces PCP induced alterations in sensorimotor gating, locomotor activity and working memory in rats [13,14,30,31]. Additionally, the acute effects of PCP administration were validated by observing an immediate increase in locomotor activity in the PCP treated pups compared to saline treated pups. Six rats from each group (PCP and control) were sacrificed at three different time points, PN12 days, 5 weeks or 14 weeks of age as described previously [18,32]. These timepoints represent perinatal, adolescent and adult developmental stages respectively, and were chosen because they are relevant and distinct brain developmental periods [33].
Rat brain tissue preparation
Rats were sacrificed by carbon dioxide asphyxiation and decapitation at PN12, 5 weeks and 14 weeks. Brains were rapidly removed and the prefrontal cortex and hippocampus was regionally dissected on ice with the aid of a standard rat brain atlas [34] Samples were snap frozen in liquid nitrogen and then stored at -80 °C. Tissue was homogenized gently in a homogenizing buffer (50 mM Tris pH 7.5, 50 % glycerol), containing a protease inhibitor cocktail (Sigma). Protein concentrations were determined by a spectrophotometer. All samples were diluted to a concentration of 2 μg/μL and stored at -80°C until required for immunoblotting, as previously detailed [35].
Immunoblotting
Relative levels of all proteins were determined by immunoblot analysis as previously described [35]. In short, proteins were resolved by SDS-PAGE with a total of 10 µg protein loaded into each well of the gel. Separated proteins were then transferred to polyvinylidene fluoride membranes (Bio-Rad). Membranes were probed with antiNWASP (1:1000; ab23394 Abcam), anti-WAVE1 (1:2500; ab50356 Abcam), anti-MBP (1:1000; ab53294 Abcam), and anti-MOG (1:10000; MAB5680 Millipore) primary polyclonal or monoclonal antibodies. Visualization and quantification of immunoblot bands was performed using the Gel Logic 2200 Pro (Carestream Molecular Imaging; Rochester, NY, USA). Samples were loaded in a randomized order with even numbers of PCP and control samples per time point per gel to minimize the effects of gel-to-gel variability on the results. A pooled sample combining aliquots from all 36 rats, was used as a positive control and was loaded onto each gel within the experiment to account for any gel-to-gel variability. Samples from each gel were then normalized to their respective pooled sample, and all immunoblot bands were normalized to a β-actin (1:5000; MAB1501 Millipore) same lane loading control. Mean β-actin expression levels did not differ between PCP and control groups (p>0.05). All experiments and quantifications were performed blind to treatment and age group.
Statistics
Statistical analyses were performed using SPSS (version 20.0, SPSS Inc. Chicago, USA). Protein analyses were performed on 5-6 biological replicates per treatment per time-point, with experiments performed in 2-3 technical replicates. As all data were normally distributed (K-S p>0.10), parametric testing was implemented. Differences between treatment and control groups were analyzed at individual time-points using unpaired two-tailed t-tests. The significance for all statistical tests was set to p<0.05
14 weeks:Perinatal PCP treatment resulted in no significant alterations in cortical levels of NWASP protein at 14 weeks during adulthood (t1,10=0.5414; p=0.601; Figure 1). Additionally there were no significant differences in WAVE1 expression in the PCP treated rats compared to controls (t1,10=0.1405; p=0.891; Figure 1). Furthermore, levels of expression of both of the tested myelination markers MBP (t1,9=0.1987; p=0.847) and MOG (t1,9=0.2912; p=0.777) were unaltered in the 14 week perinatal PCP treated rats compared to controls (Figure 1).

WAVE1 expression has been shown to be specific to the brain in mice, with the highest expression levels being detectable in the striatum, cortex and hippocampus [26]. Genetic knockout of WAVE1 results in significant reductions in hippocampal and cortical dendritic spine densities, and alterations in synaptic plasticity [25]. along with deficits in learning and memory [26]. In line with the literature, we hypothesize that the significant reduction in WAVE1 protein expression observed in our PCP treated rats, may be at least partly responsible for the synaptic and behavioural deficits observed in schizophrenia animal models involving NMDA receptor antagonism. In further support of our hypothesis, both NMDA receptor knockdown mice and PCP treated rats, in addition to WAVE1 knockout mice show similar alterations in their behavior and synaptic morphology [25,26,26,37,44,52].
Result
Prefrontal cortexPN12:Relative levels of NWASP expression were found to be significantly decreased by 23.5% in the prefrontal cortex of perinatal PCP treated rats at PN12 (t1,7=2.665; p=0.032; Figure 1). Similarly, WAVE1 expression levels were also found to be significantly decreased by 17.5% in PCP treated rats compared to control rats at PN12 (t1,7=6.117; p<0.001; Figure 1). Furthermore, there was a significant 17.5% decrease in the protein expression of the myelination marker MBP in rats treated with PCP compared to control rats at PN12 (t1,7=3.266; p=0.013; Figure 1); however there were no significant alterations in expression levels of the MOG marker of myelination (t-1,10=1.345; p=0.208) in the PN12 PCP treated rats compared to control rats (Figure 1)5 weeks:Interestingly, NWASP was the only protein to show significant alterations in expression levels in the prefrontal cortex of the adolescent 5 week old rats; and it was only a minor change with an 8% decrease in NWASP expression in the PCP treated rats at adolescence (t1,10=2.405; p=0.037; Figure 1). There were no significant alterations in the expression levels of WAVE 1(t1,10=0.4676; p=0.650), or in either of the myelination markers MBP (t1,8=0.5707; p=0.5838) or MOG (t1,10=0.6339; p=0.540) in the 5 week PCP treated rats compared to control rats (Figure 1).
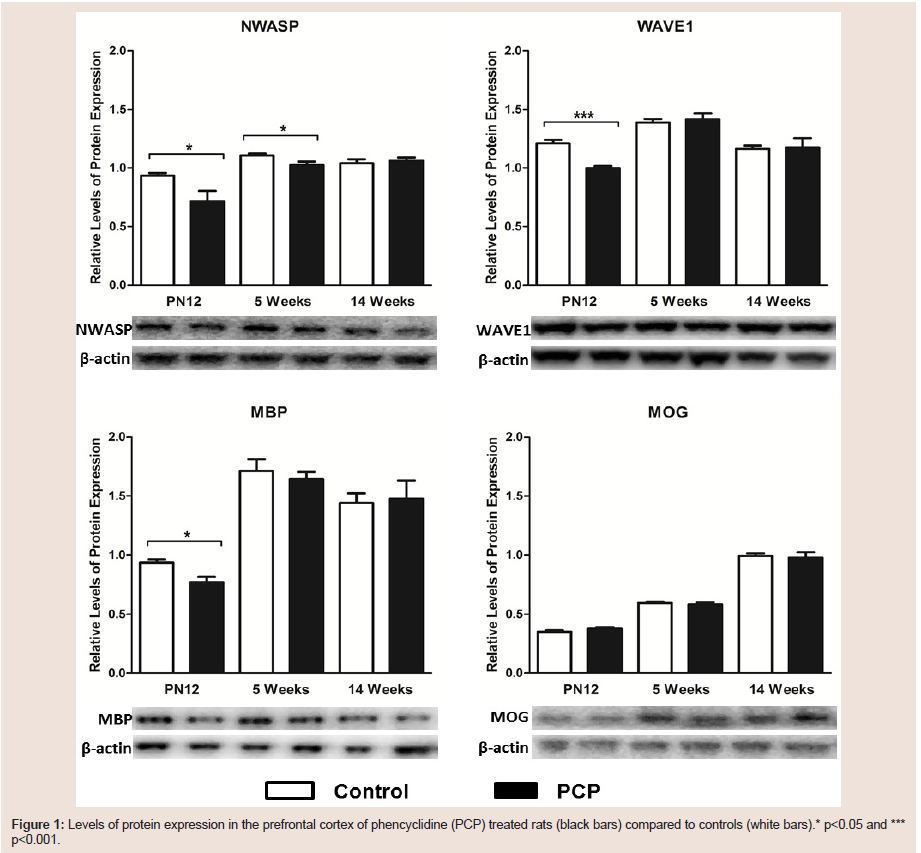
figure 1: Levels of protein expression in the prefrontal cortex of phencyclidine (PCP) treated rats (black bars) compared to controls (white bars).* p<0.05 and ***p<0.001.
Hippocampus
PN12:There was a very large 42% decrease in hippocampal WAVE1 protein expression in juvenile PCP treated rats compared to control rats (t1,10=3.659; p=0.004; Figure 2). In contrast, relative protein expression levels of NWASP (t1,10=1.456; p=0.176), and myelination markers MBP (t1,8=1.509; p=0.169) and MOG (t1/sub>,10=0.0447; p=0.965) were not significantly altered in the hippocampus of juvenile PCP treated rats (Figure 2)
5 weeks:Similarly, during adolescence, WAVE1 was the only protein to be significantly altered, with a decrease of 16% in PCP treated rats compared to controls (t1,10=3.428; p=0.006; Figure 2); whereas levels of NWASP (t1,10=0.0444; p=0.965), MBP (t1,9=1.067; p=0.314) and MOG (t1,10=0.1822; p=0.859) remained unaltered in the hippocampus of the adolescent treated rats (Figure 2).
14 weeks:Perinatal PCP treatment resulted in no significant alterations in hippocampal levels of NWASP protein during adulthood (t1,10=1.385; p=0.1962; Figure 2). Additionally there were no significant differences in adult levels of hippocampal WAVE1 expression in the PCP treated rats compared to controls (t1,10=0.8784; p=0.401; Figure 2). Furthermore, levels of expression of both of the tested myelination markers MBP (t1=0.2870; p=0.7824) and MOG (t1,9=0.7322; p=0.4827) were unaltered in the 14 week perinatal PCP treated rats compared to controls (Figure 2).
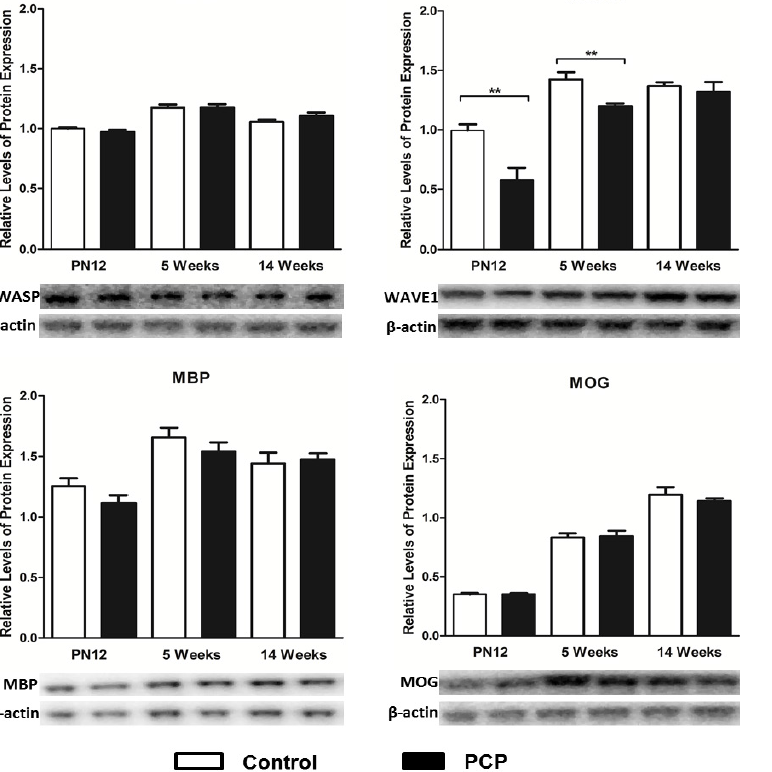
Figure 2: Levels of protein expression in the hippocampus of phencyclidine (PCP) treated rats (black bars) compared to controls (white bars). ** p<0.01.
Discussion
This is the first study to examine the developmental expression profile of the scaffolding proteins NWASP and WAVE1, in addition to myelin related proteins MBP and MOG in a neurodevelopmental PCP model of schizophrenia. This study offers insight into the role of these proteins following administration of a potent NMDA receptor antagonist during a critical neurodevelopmental period, which is also pertinent to the development of schizophrenia. In the present study, cortical levels of protein expression of NWASP, WAVE1 and MBP were significantly reduced in the juvenile PCP treated rats, and cortical levels of NWASP were slightly reduced in the adolescent PCP treated rats. In the hippocampus, WAVE1 was the only protein to have its expression levels altered, with a significant decrease observed in both PN12 and 5 week old PCP treated rats compared to their controls. Considering the current literature and the role that these scaffolding and myelin related proteins play in synaptic plasticity and myelin related processes, our results suggest that not only are NWASP and WAVE1 sensitive to the effects of NMDA receptor antagonism by PCP, but that they may contribute to the development of myelination deficits in the brain.NMDA receptor antagonism by PCP acutely reduces expression of NWASP, WAVE1 and MBP proteins in the prefrontal cortex of juvenile rats
Impaired glutamate signaling results in significant alterations in synapse number and synaptic communication, and has been correlated with neuronal, cognitive and behavioral dysfunction in a wide range of neuropsychiatric disorders including schizophrenia [36]. Similarly, PCP administration in rats has been shown to significantly reduce the number of dendritic spine synapses in the prefrontal cortex, for up to 4 weeks following the cessation of PCP treatment [37], and induce significant cognitive impairment in both monkeys [39] and rats [40-43]. NWASP and WAVE1 are important regulatory proteins involved in maintaining the actin cytoskeleton [22] and are critical for the development of dendritic spines and synapses [24,25].We have shown NWASP and WAVE1 protein expression to be significantly reduced in the prefrontal cortex of juvenile PCP treated rats compared to controls. Since neuronal changes in NWASP and WAVE1 expression have been shown to result in alterations in dendritic spine growth and synaptic transmission [24,25,27], it was not surprising to find significant reductions of both of these proteins in the prefrontal cortex of PCP treated rats. Significant deficits in NWASP and WAVE1 protein expression have previously been observed in NMDA receptor knock-down mice, with the reduced levels of the NMDA receptor having a more profound effect of on WAVE1 expression [44].
The integrity of myelin is crucial to the functionality of neurocircuitry, and disruption to myelin during development, or even after its formation can have severe neurological consequences [45]. Oligodendrocytes are known to express several regulatory actinbinding proteins including NWASP [28], and NWASP knockout PN10 neuronal cultures have been shown to have severely reduced levels of myelin related gene expression, whereby only trace amounts of MBP were detectable in the NWASP mutant cells compared to controls [46]. Moreover, WAVE1 knockout mice show defective morphogenesis of oligodendrocytes and regional hypo-myelination [29]. In addition, both NWASP and WAVE1 protein expression levels are increased at PN15, the time of onset of MBP expression in rat brain lysates [47]. To date only two studies have shown myelination deficits in rodents following PCP administration. One study administered PCP to the rats in utero [48], and the other to postnatal rat pups [49]. Both studies reported significant deficits in levels of myelination, as measured by MBP expression, in the cortex of rats exposed to PCP compared to controls [48,49]. In light of the current literature, we hypothesize that the deficits in MBP levels that we have observed in the prefrontal cortex of juvenile PCP treated rats may be caused by the significant downregulation of NWASP and WAVE1 proteins in the treated rats
Myelination processes in the hippocampus occur during the embryonic period which is a pivotal period for the development of schizophrenia [50], while myelination of the neurons within the dorsolateral prefrontal cortex occurs in late adolescence, which correlates with the age of onset of schizophrenia [51]. Despite MBP and MOG levels being unaltered in the hippocampus of PCP treated rats compared to control rats in all of the tested age groups, MBP levels as a marker of mature oligodendrocytes and myelination were significantly greater in the hippocampus than in the prefrontal cortex of the juvenile control rats (p=0.002) whereas they were not significantly different in PCP treated rats (p=0.141) (3). Furthermore although not reaching statistical significance, MBP levels were elevated in the prefrontal cortex of PCP treated compared to control rats at adolescence. These juvenile and adolescent MBP levels seen in our study are in accordance with what would be expected to occur in normal brain development, with the hippocampus being more highly myelinated during early development, and the prefrontal cortex being slow to begin with, but taking over at adolescence [50]
It is interesting that we reported differential expression patterns of myelin related protein expression across the prefrontal cortex and hippocampus following perinatal PCP treatment in this study; however it is not uncommon to observe variations in expression patterns within different regions of the brain due to the regional differences in heterogeneity in cyto- and chemo-architecture. We suspect that the perinatal PCP treatment may have affected the developmental trajectories of these myelin related proteins to varying degrees early on in development.
Hippocampal levels of WAVE1 but not NWASP or myelin related proteins MBP and MOG are altered by perinatal PCP administration
Hippocampal expression levels of WAVE1 were found to be significantly decreased in PCP treated rats compared to control rats at both juvenile and adolescent time-points, whereas hippocampal levels of NWASP were not found to be significantly altered by PCP treatment. In support of this, expression levels of WAVE1 have been shown to be more susceptible to the effects of NMDA receptor knock-down than those of NWASP [44], and WAVE1 expression is not detectable in the hippocampus of WAVE1 knockout mice [26].
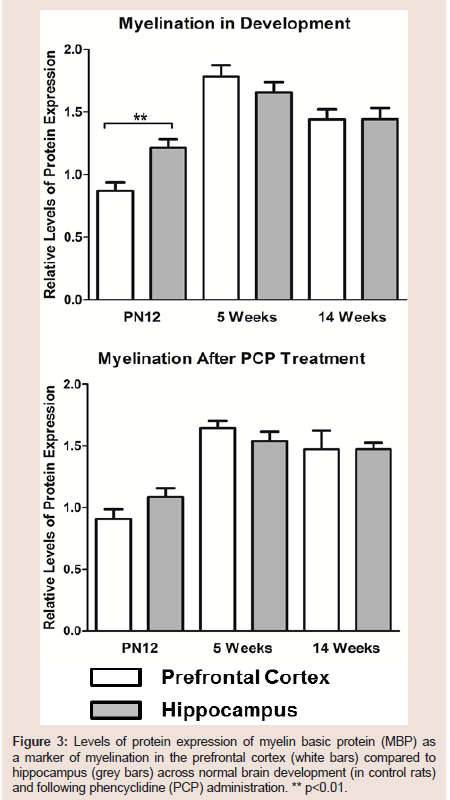
Figure 3:Levels of protein expression of myelin basic protein (MBP) as a marker of myelination n the prefrontal cortex (white bars) compared to hippocampus (grey bars) across normal brain development (in ontrol rats) and following phencyclidine (PCP) administration. ** p<0.01.
Even though the change in NWASP was very minimal across development, both NWASP and WAVE1 proteins were increased during adolescence [2]. Due to the extensive synaptic changes that occur during adolescence (reductions in dendritic arborization, and pruning of synapses) [53], it was not surprising to observe higher levels of these important regulators of the actin cytoskeleton at this critical period of development in the control rats.
Cognitive deficits observed in patients with schizophrenia are particularly attributable to prefrontal cortex and hippocampal dysfunction, and since a large number of studies have consistently found that the largest changes in the expression of oligodendrocytes and myelin-related genes in schizophrenia are in the prefrontal cortex, cingulate cortex, hippocampus and superior temporal gyrus [54-59], it seems plausible to suggest that the cognitive dysfunction seen in schizophrenia is related to the alterations in myelin-related genes in these regions. As briefly mentioned above, while we did not observe hippocampal levels of myelin related proteins to be significantly altered in PCP treated rats compared to controls, we did notice that hippocampal levels of these proteins were not significantly different from those observed in the prefrontal cortex of PCP treated rats, whereas in control rats these levels were significantly different at PN12 (Figure 3). We hypothesize that the lack of a statistically significant difference in the hippocampus compared to the prefrontal cortex levels of myelin related proteins in PCP treated rats, may be playing a role in some of the PCP induced cognitive deficits that are observed in rodent models of schizophrenia, long-term abusers of PCP [38]. and in schizophrenia patients [45]. In support of this, injections of potent demyelination agents into the ventral hippocampus of 10 day old rat pups has been shown to induce a variety of schizophrenia related endophenotypes, including deficits in prepulse inhibition, hyper locomotion and inducing anxiety related behaviors, suggesting that demyelination plays an important role in the development and severity of schizophrenia [60].
Summary and Conclusion
Here we provide the first report of alterations in the actin related cytoskeletal proteins NWASP and WAVE1, in the prefrontal cortex and hippocampus of rats from a neurodevelopmental PCP model of schizophrenia. We have shown an altered developmental trajectory of these signaling proteins following PCP administration, in particular at the perinatal stage of life. The results of this study in concert with the current literature, suggest that NWASP and especially WAVE1 are sensitive to the effects of NMDA receptor antagonism by PCP administration, and that alterations in these scaffolding related proteins may contribute to the development of myelination deficits in the brain, and more broadly, deficits in cognitive performance. Further studies will be required to fully elucidate the molecular mechanisms involved.References
- Javitt DC, Zukin SR, Heresco-Levy U, Umbricht D (2012) Has an angel shown the way? Etiological and therapeutic implications of the PCP/NMDA model of schizophrenia. Schizophr Bull 38: 958-966.
- Kapur S, Seeman P (2002) NMDA receptor antagonists ketamine and PCP have direct effects on the dopamine D-2 and serotonin 5-HT2 receptors - implications for models of schizophrenia. Mol Psychiatry 7: 837-844.
- Seeman P (2009) Glutamate and dopamine components in schizophrenia. J Psychiatry Neurosci 34: 143-149.
- Seeman P, Ko F, Tallerico T (2005) Dopamine receptor contribution to the action of PCP, LSD and ketamine psychotomimetics. Mol Psychiatry 10: 877-883.
- Amir A, Fuchs P, Gamliel A, Reis M, Shainberg A (1985) Effects of phencyclidine and analog drugs on acetylcholine receptor of cultured muscle cells. Biochem Pharmacol 34: 949-954.
- Hustveit O, Maurset A, Øye I (1995) Interaction of the chiral forms of ketamine with opioid, phencyclidine, σ and muscarinic receptors. Pharmacol Toxicol 77: 355-359.
- Maayani S, Weinstein H (1980) “Specific binding” of 3H-phencyclidine: Artifacts of the rapid filtration method. Life Sci 26: 2011-2022.
- Zukin SR (1982) Differing stereospecificities distinguish opiate receptor subtypes. Life Sci 31: 1307-1310.
- Jones C, Watson D, Fone K (2011) Animal models of schizophrenia. Br J Pharmacol 164: 1162-1194.
- Gilmour G, Dix S, Fellini L, Gastambide F, Plath N, et al. (2012) NMDA receptors, cognition and schizophrenia - Testing the validity of the NMDA receptor hypofunction hypothesis. Neuropharmacology 62: 1401-1412.
- McGonigle P (2014) Animal models of CNS disorders. Biochem Pharmacol 87: 140-149.
- Jodo E (2013) The role of the hippocampo-prefrontal cortex system in phencyclidine-induced psychosis: a model for schizophrenia. J Physiol Paris 107: 434-440.
- Du Bois TM, Huang XF, Deng C (2008) Perinatal administration of PCP alters adult behaviour in female Sprague-Dawley rats. Behav Brain Res 188: 416-419.
- Wang C, McInnis J, West JB, Bao J, Anastasio N, et al. (2003) Blockade of phencyclidine-induced cortical apoptosis and deficits in prepulse inhibition by M40403, a superoxide dismutase mimetic. J Pharmacol Exp Ther 304: 266-271.
- Harich S, Gross G, Bespalov A (2007) Stimulation of the metabotropic glutamate 2/3 receptor attenuates social novelty discrimination deficits induced by neonatal phencyclidine treatment. Psychopharmacology (Berl) 192: 511-519.
- Moghaddam B, Krystal JH (2012) Capturing the angel in “angel dust”: twenty years of translational neuroscience studies of nmda receptor antagonists in animals and humans. Schizophr Bull 38: 942-949.
- GunduzBruce H (2009) The acute effects of NMDA antagonism: From the rodent to the human brain. Brain Res Rev 60: 279-286.
- Du Bois TM, Deng C, Han M, Newell KA, Huang XF (2009) Excitatory and inhibitory neurotransmission is chronically altered following perinatal NMDA receptor blockade. Eur Neuropsychopharmacol 19: 256-265.
- Lindahl JS, Kjellsen BR, Tigert J, Miskimins R (2008) In utero PCP exposure alters oligodendrocyte differentiation and myelination in developing rat frontal cortex. Brain Res 1234: 137-147.
- Nimchinsky EA, Sabatini BL, Svoboda K (2002) Structure and function of dendritic spines. Annu Rev Physiol 64: 313-353.
- Cingolani LA, Goda Y (2008) Actin in action: the interplay between the actin cytoskeleton and synaptic efficacy. Nat Rev Neurosci 9: 344-356.
- 1. Goley ED, Welch MD (2006) The ARP2/3 complex: an actin nucleator comes of age. Nat Rev Mol Cell Biol 7: 713-726.
- Hotulainen P, Hoogenraad CC (2010) Actin in dendritic spines: connecting dynamics to function. J Cell Biol 189: 619-629.
- Wegner AM, Nebhan CA, Hu L, Majumdar D, Meier KM et al. (2008) N-wasp and the arp2/3 complex are critical regulators of actin in the development of dendritic spines and synapses. J Biol Chem 283: 15912-15920.
- Soderling SH, Guire ES, Kaech S, White J, Zhang F, et al. (2007) A WAVE-1 and WRP signaling complex regulates spine density, synaptic plasticity, and memory. J Neurosci 27: 355-365.
- Soderling SH, Langeberg LK, Soderling JA, Davee SM, Simerly R et al. (2003) Loss of WAVE-1 causes sensorimotor retardation and reduced learning and memory in mice. Proc Natl Acad Sci U S A 100: 1723-1728.
- Hering H, Sheng M (2003) Activity-dependent redistribution and essential role of cortactin in dendritic spine morphogenesis. J Neurosci 23: 11759-11769.
- Wagner JG (1973) A modern view of pharmacokinetics. J Pharmacokinet Biopharm 1: 363-401.1. Bacon C, Lakics V, Machesky L, Rumsby M (2007) N-WASP regulates extension of filopodia and processes by oligodendrocyte progenitors, oligodendrocytes, and Schwann cells-implications for axon ensheathment at myelination. Glia 55: 844-858.
- Kim HJ, DiBernardo AB, Sloane JA, Rasband MN, Solomon D (2006) WAVE1 Is Required for Oligodendrocyte Morphogenesis and Normal CNS Myelination. J Neurosci 26: 5849-5859.
- Wang C, McInnis J, Ross-Sanchez M, Shinnick-Gallagher P, Wiley JL et al. (2001) Long-term behavioral and neurodegenerative effects of perinatal phencyclidine administration: implications for schizophrenia. Neuroscience 107: 535-550.
- Wiley JL, Bühler KG, Lavecchia KL, Johnson KM (2003) Pharmacological challenge reveals long-term effects of perinatal phencyclidine on delayed spatial alternation in rats. Prog Neuropsychopharmacol Biol Psychiatry 27: 867-873.
- Du Bois TM, Newell KA, Huang XF (2012) Perinatal phencyclidine treatment alters neuregulin 1/erbB4 expression and activation in later life. Eur Neuropsychopharmacol 22: 356-363.
- Andersen SL (2003) Trajectories of brain development: point of vulnerability or window of opportunity?. Neurosci Biobehav Re 27: 3-18.
- Paxinos G, Watson C (2007) The Rat Brain in Stereotaxic Coordinates, 6th edition. Academic press.
- Fernandez Enright F, Andrews JL, Newell KA, Pantelis C, Huang XF (2014) Novel implications of Lingo-1 and its signaling partners in schizophrenia. Transl Psychiatry 4: e348.
- Penzes P, Cahill ME, Jones KA, VanLeeuwen JE, Woolfrey KM (2011) Dendritic spine pathology in neuropsychiatric disorders. Nat Neurosci 14: 285-293.
- Hajszan T, Leranth C, Roth RH (2006) Subchronic Phencyclidine treatment decreases the number of dendritic spine synapses in the rat prefrontal cortex. Biol Psychiatry 60: 639-644.
- Elsworth JD, Morrow BA, Hajszan T, Leranth C, Roth RH (2011) Phencyclidine-induced loss of asymmetric spine synapses in rodent prefrontal cortex is reversed by acute and chronic treatment with olanzapine. Neuropsychopharmacology 36: 2054-2061.
- Jentsch JD, Elsworth JD, Redmond DE, Roth RH (1997) Phencyclidine increases forebrain monoamine metabolism in rats and monkeys: Modulation by the isomers of HA966. J Neurosci 17: 1769-1775.
- Redrobe JP, Elster L, Frederiksen K, Bundgaard C, de Jong IEM, et al. (2012) Negative modulation of GABA(A) alpha 5 receptors by RO4938581 attenuates discrete sub-chronic and early postnatal phencyclidine (PCP)-induced cognitive deficits in rats. Psychopharmacology (Berl) 221: 451-468.
- Jentsch JD, Redmond DE, Elsworth JD, Taylor JR, Youngren KD (1997) Enduring cognitive deficits and cortical dopamine dysfunction in monkeys after long-term administration of phencyclidine. Science 277: 953-955.
- Horiguchi M, Hannaway KE, Adelekun AE, Jayathilake K, Meltzer HY (2012) Prevention of the phencyclidine-induced impairment in novel object recognition in female rats by Co-administration of lurasidone or tandospirone, a 5-HT 1A partial agonist. Neuropsychopharmacology 37: 2175-2183.
- Horiguchi M, Meltzer HY (2013) Blonanserin reverses the phencyclidine (PCP)-induced impairment in novel object recognition (NOR) in rats: Role of indirect 5-HT1A partial agonism. Behav Brain Res 2013 247: 158-164.
- Milenkovic M (2011) Developmental consequences of n-methyl-d-aspartate receptor hypofunction. Thesis.
- Takahashi N, Sakurai T, Davis KL, Buxbaum JD (2011) Linking oligodendrocyte and myelin dysfunction to neurocircuitry abnormalities in schizophrenia. Prog Neurobiol 93: 13-24.
- Jin F, Dong B, Georgiou J, Jiang Q, Zhang J (2011) N-WASp is required for Schwann cell cytoskeletal dynamics, normal myelin gene expression and peripheral nerve myelination. Development 138: 1329-1337.
- Wang H, Tewari A, Einheber S, Salzer JL, MelendezVasquez CV (2008) Myosin II has distinct functions in PNS and CNS myelin sheath formation. J Cell Biol 182: 1171-1184.
- Lindahl JS, Kjellsen BR, Tigert J, Miskimins R (2008) In utero PCP exposure alters oligodendrocyte differentiation and myelination in developing rat frontal cortex. Brain Res 1234: 137-147.
- Zhang R, He J, Zhu S, Zhang H, Wang H (2012) Myelination deficit in a phencyclidine-induced neurodevelopmental model of schizophrenia. Brain Res 1469: 136-143.
- Meier S, Brauer AU, Heimrich B, Nitsch R, Savaskan NE (2004) Myelination in the hippocampus during development and following lesion. Cell Mol Life Sci 61: 1082-1094.
- Weinberger DR (1987) Implications of normal brain development for the pathogenesis of schizophrenia. Arch Gen Psychiatry 44: 660-669.
- Jentsch JD, Tran A, Le D, Youngren KD, Roth RH (1997) Subchronic phencyclidine administration reduces mesoprefrontal dopamine utilization and impairs prefrontal cortical-dependent cognition in the rat. Neuropsychopharmacology 17: 92-99.
- Sisk CL, Zehr JL, (2005) Pubertal hormones organize the adolescent brain and behavior. Front Neuroendocrinol 26: 163-174.
- Aston C, Jiang L, Sokolov BP (2005) Transcriptional profiling reveals evidence for signaling and oligodendroglial abnormalities in the temporal cortex from patients with major depressive disorder. Mol Psychiatry 10: 309-322.
- Aston C, Jiang L, Sokolov BP (2004) Microarray analysis of postmortem temporal cortex from patients with schizophrenia. J Neurosci Res 77: 858-866.
- Dracheva S, Davis KL, Chin B, Woo DA, Schmeidler J (2006) Myelin-associated mRNA and protein expression deficits in the anterior cingulate cortex and hippocampus in elderly schizophrenia patients. Neurobiol Dis 21: 531-540.
- Katsel P, Davis KL, Haroutunian V (2005) Variations in myelin and oligodendrocyte-related gene expression across multiple brain regions in schizophrenia: A gene ontology study. Schizophr Res 79: 157-173.
- Tkachev D, Mimmack ML, Ryan MM, Wayl M, Freeman T, et al. (2003) Oligodendrocyte dysfunction in schizophrenia and bipolar disorder. Lancet 362: 798-805.
- Hakak Y, Walker JR, Li C, Wong WH, Davis KL, et al. (2001) Genome-wide expression analysis reveals dysregulation of myelination-related genes in chronic schizophrenia. Proc Natl Acad Sci U S A 98: 4746-4751.
- Makinodan M, Yamauchi T, Tatsumi K, Okuda H, Takeda T, et al. (2009) Demyelination in the juvenile period, but not in adulthood, leads to long-lasting cognitive impairment and deficient social interaction in mice. Prog Neuropsychopharmacol Biol Psychiatry 33: 978-985.