
Journal of Oral Biology
Download PDF
Research Article
*Address for Correspondence: Daniel H. Fine, Professor and Chairman of the Department of Oral Biology and Director of the Center for Oral Infectious Diseases from Rutgers University, 185 South Orange Avenue, Medical Science Building, Room C-636, Rutgers School of Dental Medicine, Newark, New Jersey 07103, USA, Tel: 973-972-3728; Fax: 973-972-0045; E-mail: finedh@sdm.rutgers.edu
Citation: Fine DH, Karched M, Furgang D, Sampathkumar V, Velusamy S, et al. Colonization and Persistence of Labeled and “Foreign” Strains of Aggregatibacter actinomycetemcomitans Inoculated into the Mouths of Rhesus Monkeys. J Oral Bio. 2015;2(1): 10.
Copyright © 2015 Fine et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Oral Biology | ISSN: 2377-987X | Volume: 2, Issue: 1
Submission: 19 December 2014 | Accepted: 23 January 2015 | Published: 27 January 2015

Initially we screened over 80 Macaca mulatta (Rhesus; Rh) monkeys, 40 marmosets, and an assortment of other primates including; Chimpanzees (n=8), Cynomolgus (n=4), Baboons (n=4), and Tamarins (n=4) from three primate centers to determine the percentage of primates who harbored A. actinomycetemcomitans. A. actinomycetemcomitans was isolated from both buccal tissue and plaque obtained from each of the 140 primates screened. These samples were then grown on AAGMBV agar and then specific colonies with the characteristic colonial morphology were picked from plates and tested using biochemical and DNA methods for identification. We then subcultured several A. actinomycetemcomitans stains and extracted DNA to do analysis of selected virulence traits for our initial comparative genomic assessment [7]. More than 80% of the Rh monkeys sampled had A. actinomycetemcomitans in their dental plaque [20]. Our goal was to introduce a labeled stain of A. actinomycetemcomitans into a complex competitive environment that was capable of supporting its growth and survival.

Buccal epithelial cell (BEC) binding assay
 Selection of human and monkey A. actinomycetemcomitans strains for study
Selection of human and monkey A. actinomycetemcomitans strains for study



Choice of food for repeated inoculation after cleaning
 Preparation for inoculation: debridement, feeding and inoculation methods
Preparation for inoculation: debridement, feeding and inoculation methods

Colonization and Persistence of Labeled and “Foreign” Strains of Aggregatibacter actinomycetemcomitans Inoculated into the Mouths of Rhesus Monkeys
Daniel H. Fine1*, Maribasappa Karched2, David Furgang1, Vandana Sampathkumar1, Senthil Velusamy1 and Dipti Godboley1
- 1Department of Oral Biology, Rutgers School of Dental Medicine, Rutgers University, USA
- 2Faculty of Dentistry, Bioclinical Sciences, Kuwait University, USA
*Address for Correspondence: Daniel H. Fine, Professor and Chairman of the Department of Oral Biology and Director of the Center for Oral Infectious Diseases from Rutgers University, 185 South Orange Avenue, Medical Science Building, Room C-636, Rutgers School of Dental Medicine, Newark, New Jersey 07103, USA, Tel: 973-972-3728; Fax: 973-972-0045; E-mail: finedh@sdm.rutgers.edu
Citation: Fine DH, Karched M, Furgang D, Sampathkumar V, Velusamy S, et al. Colonization and Persistence of Labeled and “Foreign” Strains of Aggregatibacter actinomycetemcomitans Inoculated into the Mouths of Rhesus Monkeys. J Oral Bio. 2015;2(1): 10.
Copyright © 2015 Fine et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Oral Biology | ISSN: 2377-987X | Volume: 2, Issue: 1
Submission: 19 December 2014 | Accepted: 23 January 2015 | Published: 27 January 2015
Abstract
Aggregatibacter actinomycetemcomitans (Aa) is a pathobiont and part of a consortium of bacteria that can lead to periodontitis in humans. Our aim was to develop a model for oral inoculation of labeled Aa into a suitable host in order to study Aa traits and ecological factors that either enhance or repress its persistence.Primate species were screened for Aa to select a host for colonization studies. Macaca mulatta (Rhesus/Rh) was selected. RhAa strains were isolated, subjected to sequencing and functional analysis for comparison to human strains. “Best” methods for microbial decontamination prior to inoculation were assessed. Three groups were studied; Group 1 (N=5) was inoculated with Aa Spectinomycin resistant (SpecR) Rh strain 4.35, Group 2 (N=5) inoculated with Aa SpecR human strain IDH 781, and Group 3 (N=5) the un-inoculated control. Repeated feeding with pancakes spiked with SpecR Aa followed high dose oral inoculation. Cheek, tongue, and plaque samples collected at baseline 1, 2, 3, and 4 weeks after inoculation were plated on agar; 1) selective for Aa, 2) enriched for total counts, and 3) containing 50 μg/ml of Spec. Aa was identified by colonial morphology and DNA analysis.
Rh and human Aa had > 93-98% genome identity. Rh Aa attached to tissues better than IDH 781 in vitro (p < 0.05). SpecR IDH 781 was not recovered from any tissue at any time; whereas, Rh SpecR 4.35 was detected in plaque, but never tongue or cheek, in all monkeys at all times (> 1 x 105 colonies/ml; p < 0.001).
In conclusion, the primate model provides a useful platform for studying integration of Aa strains into a reduced but established oral habitat. Primate derived SpecRAa was consistently detected in plaque at all collection periods; however, human derived Aa was never detected. The model demonstrated both microbial as well as tissue specificity.
Keywords
Aggregatibacter actinomycetemcomitans; Biofilms; Dysbiosis; Experimental animal models; Macaca mulatta; Periodontitis; Microbial interactionsAbbreviations
Aa: Aggregatibacter actinomycetemcomitans; Rh: Rhesus; HACEK: Haemophilus, Actinobacillus, Cardiobacteria, Eikenella, Kingella; BECs: Buccal Epithelial Cells; SHA: Salivary Coated Hydroxyapatite; AAGM: Actinobacillus Actinomycetemcomitans Growth Media; PBS: Phosphate Buffered Saline; SpecR: Spectinomycin resistance; CFU’s: Colony Forming Units; NEPRC: New England Primate Research CenterIntroduction
Aggregatibacter (formerly known as Actinobacillus) actinomycetemcomitans is a Gram-negative bacterium that colonizes the oral cavity of man and primates [1]. This capnophilic bacterium has been highly associated with localized aggressive periodontitis in individuals of African descent [2]. A. actinomycetemcomitans is a member of the HACEK (Haemophilus, Actinobacillus, Cardiobacterium, Eikenella, Kingella) group of bacteria that are also associated with a number of non-oral but systemic infections that include; coronary, kidney, brain, lung and joint diseases in man [3]. All of the HACEK bacteria but particularly Haemophili, Eikenella, and Aggregatibacter are found in dental plaque and have been detected on or in oral mucosa and can disseminate via the bloodstream [2]. Therefore, the oral cavity has been portrayed as the portal of entry for these organisms [3,4].The research focus of our group has been on A. actinomycetemcomitans associated host/microbial interactions leading to periodontal bone loss [5]. Along these lines our primary interest is in understanding how A. actinomycetemcomitans colonizes teeth, migrates below the gum-line, and avoids immune elimination [6]. Towards these ends our group has developed both in vitro assays and in vivo animal models to study A. actinomycetemcomitans colonization and survival [7,8]. Most recently, we demonstrated that A. actinomycetemcomitans leukotoxin, previously thought to be active exclusively in humans and primates, is a prominent virulence factor associated with bone resorption in a rat model of periodontal disease [9].
While we have had significant success in studying colonization in our rat model, the in vitro rat lymphocytic response to A. actinomycetemcomitans leukotoxin was modest suggesting that the in vivo rat model under-represented the bone loss detected [9]. Furthermore bone loss is the end stage of disease [10]. Moreover, there is little doubt that the primates provide a more accurate model of colonization and disease when compared to rodents [11]. For example, A. actinomycetemcomitans adhesins and leukotoxin have been shown to demonstrate similar levels of specificity with respect to human and primate tissue as compared to rodent tissue [12,13]. The main goal of this study is to develop a system that will permit us to implant A. actinomycetemcomitans into its “natural” environment such that we can study host and microbial interactions with the newly implanted species overtime. Success in this endeavor will allow us to examine how A. actinomycetemcomitans integrates into an existing but reduced oral habitat.
The integration of a “newly” implanted pathobiont into a relatively stable biofilm community can be considered as an important early event in disease development [14,15]. Previously, we have shown that A. actinomycetemcomitans can be an early tooth colonizer by demonstrating that it can be detected on an initially sterile hydroxyapatite tooth surrogate within 4-6 hours following placement into the mouths of individuals who harbor A. actinomycetemcomitans [16]. This in vivo tooth colonization by A. actinomycetemcomitans has been shown earlier in several old world primate studies but results from these experiments have not been fully appreciated and as a result A. actinomycetemcomitans is still considered by many to be a poor tooth colonizer [17-19]. Further documentation of early colonization by A. actinomycetemcomitans in humans is problematic because; 1) A. actinomycetemcomitans is not typically detected as a prominent member of the indigenous oral flora in periodontally healthy individuals, 2) the percentage of individuals colonized by A. actinomycetemcomitans is low, and 3) very little flexibility can be tolerated with respect to either microbial or environmental manipulation when humans are involved [3].
It is our contention that we can overcome these impediments by studying A. actinomycetemcomitans colonization in a mammalian environment similar to humans that is also hospitable for A. actinomycetemcomitans growth and survival. Such an environment is routinely found in primates [20]. Therefore, a primate model is appealing for the following reasons; 1) the oral environment supports the growth of A. actinomycetemcomitans, 2) microbial genes of implanted A. actinomycetemcomitans strains can be manipulated, 3) interaction of the resident flora can be studied and manipulated, and 4) host interactions can be studied and altered by factors such as smoking, antibiotics, hormones, foods etc.
Thus a model permitting the integration of a “foreign”, labeled, and newly implanted bacterium into a complex biofilm community could help address several questions. For example, tracking an implanted and labeled reference strain of A. actinomycetemcomitans can help describe both disruption and/or integration into an existing complex, compact microbial community [21,22]. This is a compelling line of investigation since all mucosal infections occur in a competitive environment and disruption or dysbiosis of that community forms the basis of the first step in the initiation of bacterially induced mucosal infections [3]. In this manuscript we describe the procedures and results derived from our studies that were designed to determine the best method and most appropriated A. actinomycetemcomitans strain capable of integrating into a microbial community that has the ability to support A. actinomycetemcomitans colonization, growth and survival, and/or elimination.
Materials and Methods
Survey of primate oral flora and selection of A. actinomycetemcomitans strains for use in the primate modelThe Institutional Animal Care and Use Committees (IACUC) at Rutgers, the New England Primate Research Center (NEPRC) and the Southwest Primate Center approved all work with primates. In all experiments, prior to anesthetizing primates, vital signs and weights were obtained. Monkeys were monitored on a daily basis as per IACUC recommendations. In cases where “foreign” strains of A. actinomycetemcomitans were implanted; procedures were performed at the conclusion of experiments to assure that these strains were no longer detectable (see Figure 1).

Figure 1: Flow Diagram illustrating the overall inoculation, colonization and persistence protocol Bacteria were collected from teeth at baseline. Animals were divided into three groups. After debridement and decontamination, inoculation was performed followed by pancake feeding on days 2 to 5. Samples were collected on weeks 1 to 3 using a buccal brush passed over one quadrant. A brush was passed over the cheeks and tongue. Brush samples were stabbed into an agar containing tube, shipped to Rutgers and samples were then plated on various media. On week 4 (28 days) samples were collected from all surfaces by scaler and vigorously with the buccal brush for plating. On day 33 decontamination was performed and on day 60 samples were collected by buccal brush and shipped for analysis of residual labeled Aa. One of the controls was dropped due to medical complications unrelated to the study. Thus 4 controls completed the study.
Of the primates screened, Rh monkeys appeared to provide the best environment (see Table 1) for our experimental goals [20]. We collected several dozen A. actinomycetemcomitans strains with a range of serotypes from the Rh primates at the NEPRC (Table 1). We compared the attachment characteristics of several of these primate strains to human A. actinomycetemcomitans strain IDH 781, considered an outstanding colonizing bacterium in vitro and in our rat studies [9,13]. Initially we compared binding of various A. actinomycetemcomitans primate and human strains to buccal epithelial cells (BECs) and salivary-coated hydroxyapatite (SHA). Several primate strains were sequenced and compared both for their complete genomes and also for a variety of virulence genes of interest [3,4]. For purposes of this study we focused our comparison of A. actinmycetemcomitans strains on attachment to soft and hard tissue since our goal was colonization.
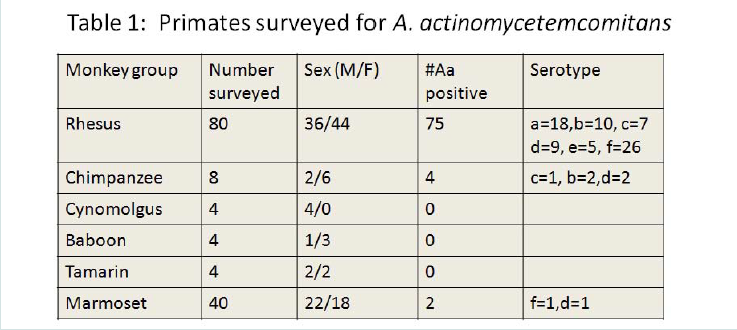
Table 1: Primates surveyed for A. actinomycetemcomitans
This assay has been described previously [7]. Briefly, we collected buccal cells from Rh monkeys by scraping the side of the cheek gently but firmly to capture enough cells to obtain 5 x 104 BECs per ml. We suspended these cells in phosphate buffered saline (PBS) and added A. actinomycetemcomitans cells (either Rh or human strains) at a concentration of 1 x 108 /ml. Prior to mixing, the A. actinomycetemcomitans suspension was subjected to sonication for 15 seconds at a low power output to break up clumps of cells.
The mixture was allowed to incubate for 30 minutes to allow for binding of A. actinomycetemcomitans to BECs. The mixture of A. actinomycetemcomitans cells and BECs was then placed over a tube containing a 5% Ficoll 400 gradient, which was subjected to low speed centrifugation (50-x g for 5 min) to separate the heavier epithelial cells from the bacteria. The supernatant was removed and the pellet containing the BECs was plated on a medium containing AAGM agar with Bacitracin and Vancomycin (AAGMBV) to determine the number of A. actinomycetemcomitans bound per BEC [16]. Primate strains RHAA, Rh 1.4, Rh 2.3, Rh 3.8, Rh 4.35 and human strain IDH 781 were used in binding studies. Controls consisted of A. actinomycetemcomitans alone, BECs alone, and BECs mixed with non-adherent lab strains of A. actinomycetemcomitans.
Binding to hydroxyapatite (HA)
This assay has also been described previously [7,23]. Briefly, a suspension of A. actinomycetemcomitans (both human and Rh derived) was obtained as described above to achieve an optical density A560 = 0.9 (equivalent to 1 x 108 cells/ml) [7]. Whole unstimulated saliva was collected from healthy subjects (an IRB from Rutgers University was used for consent) and clarified by centrifugation at 10,000-x g to obtain 1 ml of saliva for addition to 50 mg of hydroxyapatite beads (BDH Chemicals Ltd. Poole, UK). After 1 hour, beads were washed three times with KCl (0.05 mM KCl, 1 mM potassium phosphate, pH 6.0, 1 mM calcium chloride, 0.1 mM magnesium chloride) (Sigma). For attachment studies, stocks of human IDH 781 and Rh 4.35 A. actinomycetemcomitans were grown for 24 hours in AAGM broth. A 100 μl sample of the suspension was added to 50 mg of saliva coated beads and the mixture of beads and bacteria were placed on a rotating device (RotoTorque from Cole Palmer Instruments, Chicago IL) for 30 minutes. Beads were allowed to settle for 2 minutes and the supernatant containing the unbound bacteria were removed and plated on AAGM agar. The pellet containing the cells bound to SHA was then washed three times in phosphate buffered saline (PBS). After the final wash, cells were sonicated, centrifuged at low speed and serially diluted, resuspended in 1 ml of PBS, and plated on AAGM agar to determine A. actinomycetemcomitans cells bound to SHA. The ratio of bound to unbound cells was calculated as described [7,23]. Binding of primate strain Rh 4.35 was compared to human strain IDH 781.
Competitive binding to BECs
Attachment of A. actinomycetemcomitans to BECs is dependent on specific adhesin/receptor interactions that; 1) require affinity, and 2) reach a concentration dependent binding plateau. Reversibility of Rh 3.8 A. actinomycetemcomitans binding to BECs was challenged by other strains of A. actinomycetemcomitans derived from Rh monkeys (Rh 4.35) and human A. actinomycetemcomitans (IDH 781). The experiment was designed to determine if one strain of monkey Rh (4.35 SpecR) could replace the already bound Rh A. actinomycetemcomitans (Rh 3.8) in vitro in comparison to a human SpecR A. actinomycetemcomitans (IDH 781). We thought these results might serve as a possible prelude to what might happen in the mouths of monkeys. Specifically, after an initial BEC binding experiment where Rh BECs were treated with unlabeled Rh 3.8 A. actinomycetemcomitans for a 30-minute incubation period, the reaction mixture (Rh A. actinomycetemcomitans plus Rh BECs) was divided into three equal aliquots of 500 ml. One aliquot was used as the control and reacted with 500 μl of the same Rh 3.8 strain, one was reacted with 500 μl of SpecR labeled Rh A. actinomycetemcomitans (see labeling method below) derived from a different monkey strain (Rh 4.35) and the third was reacted with 500 μl of IDH 781 SpecR A. actinomycetemcomitans. All three mixtures were then incubated for another 30 minutes. After this second incubation period each reaction mixture was placed over a Ficoll gradient in preparation for assessment of SpecR A. actinomycetemcomitans binding per BEC. The goal was to determine the ability of added and labeled A. actinomycetemcomitans (human or primate) to displace the original primate A. actinomycetemcomitans (Rh 3.8) that was already bound to primate BECs [24].
Labeling of primate and human A. actinomycetemcomitans strains
IDH 781 and Rh 4.35 were labeled by creating Spectinomycin resistance (SpecR). Spectinomycin was recommended as the antibiotic of choice for labeling by the veterinarians via the IACUC because this antibiotic was not needed for treatment or other purposes. Labeling (resistance) was developed by passaging the strains on successive plates of agar containing increasing amounts of Spectinomycin until they demonstrated a level of resistance to 50 μg/ml of Spectinomycin. After successful labeling was attained, binding to BECs and SHA was repeated to determine whether the labeling had any effect on these important phenotypes.
Selection of an oral decontamination method
In preliminary experiments we were able to assess three different cleaning methods to reduce the biofilm that existed on the teeth prior to inoculation of the test A. actinomycetemcomitans strains using a subset of three primates per experiment. The first method consisted of scaling all teeth using a typical manual scaling procedure followed by inoculation of the labeled A. actinomycetemcomitans test strain. The second method included scaling, followed by rubber cup prophylaxis prior to inoculation. The third method consisted of scaling, rubber cup prophylaxis, followed by topical application of a local antiseptic (Listerine antiseptic) followed by rinsing with sterile water to remove the residual antiseptic prior to inoculation.
Selection of inoculation and feeding method
After cleaning and local inoculation of A. actinomycetemcomitans we fed the animals repeatedly to continue exposure to the inocula for the next 4 days. Both primate and human A. actinomycetemcomitans strains were first tested in vitro by adding A. actinomycetemcomitans to various foods to determine the best food substance to use for repeated inoculation so as to avoid the need to anesthetize the animals on re-inoculation. The foods assessed were banana, marshmallow, graham crackers, and pancakes. To test survival, one ml of a dose of 1 x 108 cells per ml were added to each food source and the A. actinomycetemcomitans was allowed to sit in the food source at room temperature for 0 minutes, 30 minutes and 120 minutes. Foods were then mashed and diluted in AAGM broth and tested for growth on AAGMBV agar cultured at 37 °C for three days and then A. actinomycetemcomitans colony forming units per ml (CFU’s/ml) were counted.
Selection of an inoculation method
To determine the best method of inoculation we tested feeding alone vs. direct inoculation and feeding.
Experimental Design
The purpose of the initial full experimental protocol was to determine whether a human A. actinomycetemcomitans strain or a primate A. actinomycetemcomitans strain would colonize the oral cavity of primates and survive for a period of 4 weeks. (Figure 1; Flow chart). Three groups of monkeys were used as follows; Group 1 had 5 Rh monkeys who were inoculated with SpecR labeled human strain IDH 781, Group 2 had 5 Rh monkeys who were inoculated with SpecR labeled primate strain Rh 4.35 serotype b, and Group 3 consisted of 5 Rh monkeys that were un-inoculated. Monkeys were anesthetized with 10-20 mg/kg of ketamine and medetomidine intramuscularly, brought to the laboratory suite, and then each monkey was sampled for total bacteria and recovery of indigenous A. actinomycetemcomitans. Two independent samples were taken from teeth, tongue and cheek; one for DNA analysis and one for cultural analysis of A. actinomycetemcomitans indigenous to the oral cavity of each of the primates participating in the study. Following this teeth were cleaned with scalers and then polished with a rubber cup and pumice. After polishing, teeth were washed with water, followed by application of fully concentrated Listerine to all teeth and soft tissue surfaces by means of a soaked cotton applicator. Sterile water was then applied to all surfaces to remove the Listerine. Shortly thereafter a confluent suspension of the A. actinomycetemcomitans strains (IDH 781SpecR or Rh 4.35 SpecR) was applied to all teeth and tissue surfaces until the full 1 ml of a concentration of 1 x 108 cells/ml was inoculated. Prior to inoculation the dense suspension was subjected to agitation to break up clumps. Teeth and tissue were doused by repeated topical application of a total of 1 ml of the fully dispersed thick suspension of bacteria applied to teeth and soft tissues. To subject animals to repeated inoculation, pancakes (Aunt Jemima) were then spiked with A. actinomycetemcomitans on one side and with pancake syrup on the opposite side for feeding of primates on days 2-5. Pancake fed to control animals had no A. actinomycetemcomitans.
Sampling and identification of A. actinomycetemcomitans
Animals were anesthetized and sampled as described at weeks 1, 2, and 3 using a buccal brush collection method. In this method the brush was passed over a limited area of tissue or tooth several times to collect sufficient material. Individual brushes were used to obtain samples from tooth, cheek, and tongue. After collection, brushes were stabbed into AAGMBV agar for recovery of A. actinomycetemcomitans. Samples were shipped overnight from the NEPRC to Rutgers. Samples recovered from stabs were plated on AAGMBV, Spec and enriched trypicase soy agar (ETSA) for analysis. A. actinomycetemcomitans was identified by; colonial morphology, biochemical testing, and DNA analysis [25]. Plaque samples were removed from teeth by dental scalers in the week 4-collection period (day 28), while tissue was sampled by buccal brush. This sampling was vigorous and thorough. On day 33 full prophylaxis and decontamination was performed to eliminate the inoculated SpecR A. actinomycetemcomitans (Figure 1). A final sample was taken on day 60 to insure that the inoculated A. actinomycetemcomitans strains were eliminated.
Microbiological analysis
Two samples were taken from each of the oral surfaces; one for DNA analysis, and one for cultural analysis of CFU’s/ml. Samples for DNA analysis were refrigerated and then frozen for future study. Each sample taken for cultural analysis was place in 1 ml of PBS and then plated on the following agar; ETSA with 5% blood, AAGMBV agar for enumeration of total A. actinomycetemcomitans and AAGM with 50 μg/ ml Spec for enumeration of SpecR strains of A. actinomycetemcomitans [7,20,26].
Statistical analysis
Selection of the best method for preparation of the oral cavity, for A. actinomycetemcomitans inoculation, and feeding was descriptive in nature and based on a small subset of monkeys and thus determined by inspection of data by members of the research team. Survey of the presence of A. actinomycetemcomitans in the mouths of the various primates analyzed from the three primate centers was presented in tabular form as descriptive data with no statistical analysis although it was clear without statistical analysis that Rh primates had the highest carriage of A. actinomycetemcomitans.
Data derived from in vitro binding assays (BEC) were analyzed by ANOVA using a Fisher exact test to determine the level of binding and using a Chi-square to determine best binding /when comparing the human and monkey strains to attain a significance level of 0.05. SHA binding was assessed by means of a two-way students-t-test setting the significance level at 0.05.
Data derived from the full experimental in vivo protocol designed to assess the presence or absence of SpecR A. actinomycetemcomitans in the primate samples was analyzed by ANOVA using the Kruskal Wallis analysis of variance by ranks to compare the three groups and to determine whether human or primate A. actinomycetemcomitans was better in colonizing in vivo. The un-inoculated group acted as the control in the analysis with the significance value set at p<0.05.
Results
Survey of primates for possession of A. actinomycetemcomitansIn Table 1 we present the data from our survey of over 80 Rhesus monkeys, 40 marmosets, 8 chimpanzees and 4 cynomolgus, 4 Baboons, and 4 Tamarins. While 57% of the primates sampled had A. actinomycetemcomitans, over 90 % of the Rh monkeys surveyed had A. actinomycetemcomitans. Moreover, all major serotypes of A. actinomycetemcomitans in the Rh primates were detected and 80% of the Rh primates screened had only one serotype of A. actinomycetemcomitans. Fifty percent of the chimpanzees had A. actinomycetemcomitans and only 5% of the marmosets had A. actinomycetemcomitans. We could not recover A. actinomycetemcomitans strains from the other primates evaluated (Table 1). Serotype distribution was seen as follows; “a”, and “f” were dominant but “b”, “d” and “c” were numerous (Table 1). It should be pointed out that none of the isolated strains showed the JP2 promoter region deletion.
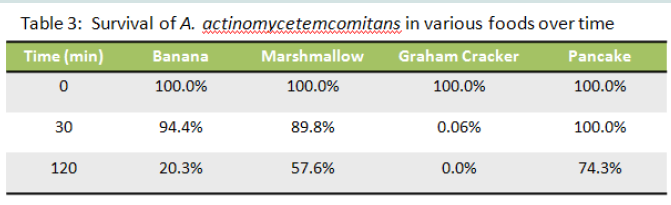
Table 1: Survival of A. actinomycetemcomitans in various foods overtime
Based on our survey and pragmatic considerations we decided to use Rh monkeys from the NEPRC for our study and initially chose a serotype b strain of A. actinomycetemcomitans (Rh 4.35) for inoculation. We performed an overall genome sequence analysis of “a”, “b”, “c” and “d” serotypes from A. actinomycetemcomitans strains isolated from Rh monkeys from the NEPRC primate center [27]. We also chose IDH 781, a human “d” serotype A. actinomycetemcomitans strain for our comparison in the inoculation studies because of its success in our in vitro binding assays and in our in vivo rat studies [9].
DNA comparison and SHA and BEC binding comparisons
After strain selection we performed DNA analysis of the genomes of eight primate strains as well as DNA and amino acid comparison of the specific virulence traits of these strains to those found in humans [27]. As such we compared human and primate strains for adhesin and attachment genes aae, apiA, flp and tadA, other virulence genes were also compared as well as an assortment of housekeeping genes. Initial analysis indicated that strains assessed had a minimum of 98% nucleic acid identity and from 93-99% sequence identity in all virulence traits analyzed suggesting that the microbes were comparable (data not shown). Since we felt that attachment characteristics were clinically relevant for our in vivo colonization experiments we compared binding properties in several primate strains and compared these properties to IDH 781 for both soft and hard tissue binding. In BEC binding no statistical differences were seen when most Rh strains were compared although strains Rh 4.35 bound better than RHAA and Rh 2.3 (p < 0.05) while comparisons to other Rh strains were not significant (Figure 2). Further in a competitive soft tissue-binding assay it was shown that Rh 4.35 monkey derived A. actinomycetemcomitans displaced Rh 3.8 monkey A. actinomycetemcomitans bound to monkey BECs by 69.8% (Table 2). These results were significantly superior to the displacement of bound Rh 3.8 A. actinomycetemcomitans by human IDH 781 A. actinomycetemcomitans which was at a level of 50.2% p > 0.05; (Table 2). Further in SHA binding comparisons bound to unbound ratios were significantly higher in primate strain Rh 4.35 (3.88 + 0.45) as compared to human strain IDH 781 (1.98 + 0.25) (Figure 3, p < 0.05). We also compared binding of the SpecR strains to their parent unlabeled strains for both hard and soft tissue binding. No significant differences were seen when labeled strains were compared to unlabeled strains (data not shown).
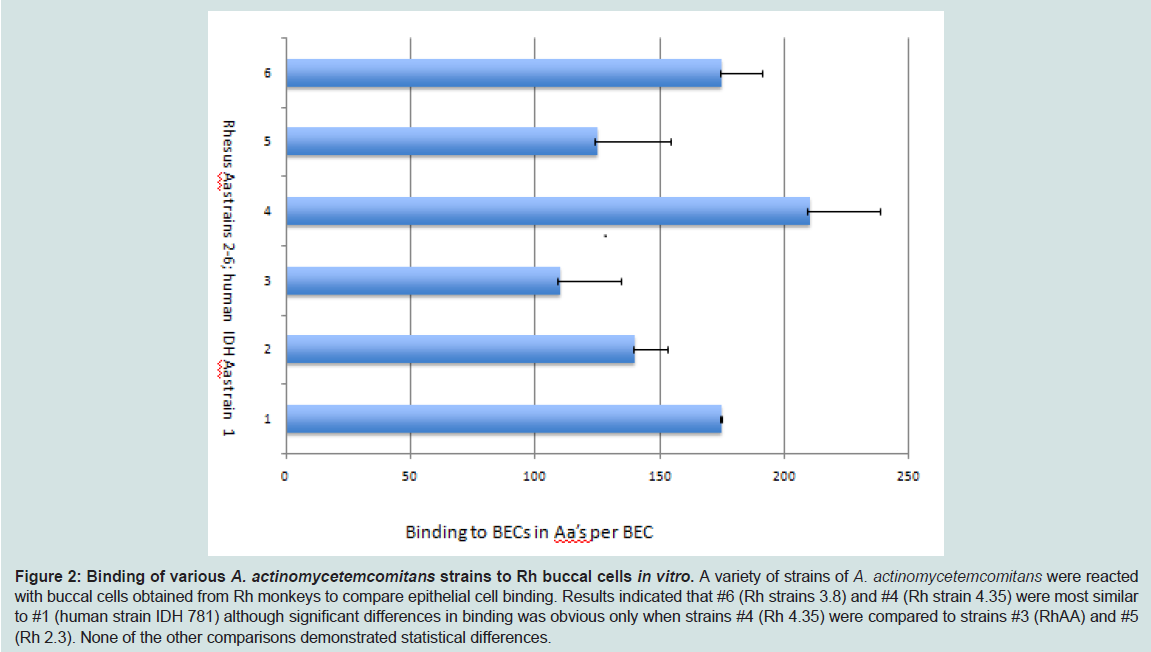
Figure 2: Binding of various A. actinomycetemcomitans strains to Rh buccal cells in vitro. A variety of strains of A. actinomycetemcomitans were reacted with buccal cells obtained from Rh monkeys to compare epithelial cell binding. Results indicated that #6 (Rh strains 3.8) and #4 (Rh strain 4.35) were most similar to #1 (human strain IDH 781) although significant differences in binding was obvious only when strains #4 (Rh 4.35) were compared to strains #3 (RhAA) and #5 (Rh 2.3). None of the other comparisons demonstrated statistical differences.

Table 2: Repalcement of Rhesus A. actinomycetemcomitans bound to Buccal Epithelial Cells(BECs).
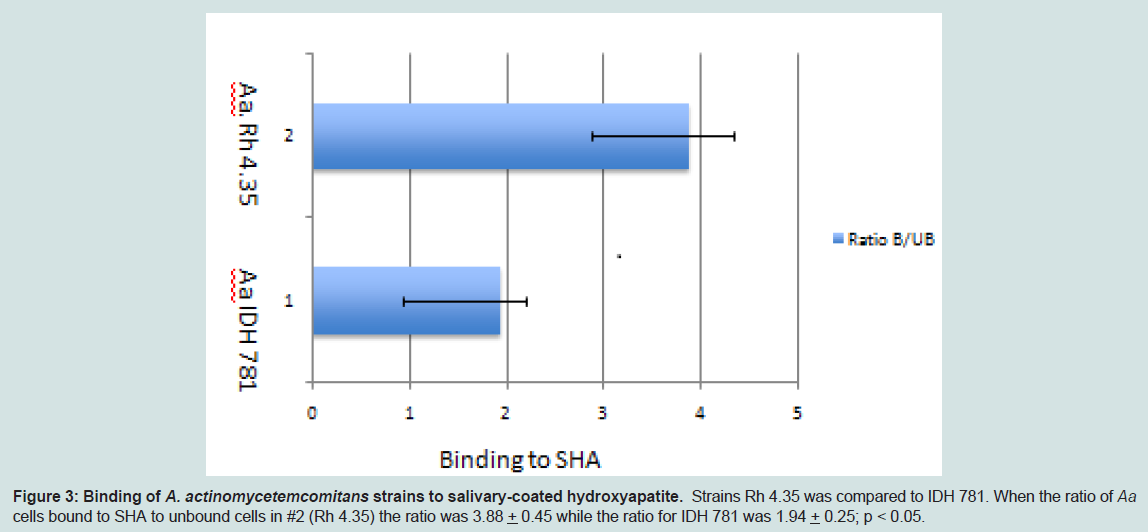
Figure 3: Binding of A. actinomycetemcomitans strains to salivary-coated hydroxyapatite. Strains Rh 4.35 was compared to IDH 781. When the ratio of Aa cells bound to SHA to unbound cells in #2 (Rh 4.35) the ratio was 3.88 + 0.45 while the ratio for IDH 781 was 1.94 + 0.25; p < 0.05.
As seen in Table 3 comparison of survival of A. actinomycetemcomitans when placed on banana, marshmallow, graham crackers and pancakes showed that A. actinomycetemcomitans survival overtime was superior on pancakes and thus 74% of A.actinomycetemcomitans were still viable after 2 hours. Prior to these in vitro experiments we gave monkeys each of the food substances as recommended by the veterinarians because the animals are known to be very particular about the form, shape, and smell of the foods they eat. To make the A. actinomycetemcomitans spiked pancakes more appealing we placed syrup on the side of the pancake free of the inoculum. This approach was well accepted by all the monkeys in the study.
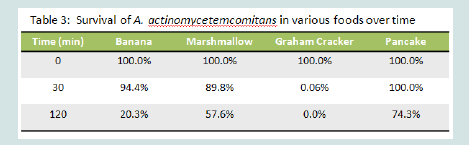
Table 3: Repalcement of Rhesus A. actinomycetemcomitans bound to Buccal Epithelial Cells(BECs).
In pilot studies we compared three forms of debridement and the method that combined manual scaling, rubber cup prophylaxis with prophylaxis paste, and topical application of Listerine, and immediate rinsing with water, provided the best preparatorymethod prior to inoculation of the “foreign” antibiotic labeled A. actinomycetemcomitans (data not shown).
In the initial pilot studies we used human strain IDH 781 SpecR as the inoculum and compared implantation and feeding methods that consisted of topical inoculation alone or in combination with feeding procedures that involved A. actinomycetemcomitans spiked pancakesIn these pilot studies we assessed survival over a two-week period following debridement. Our data indicated that only the inoculation and repeated feeding protocol was successful but colonization of the teeth was very sparse and occurred in only one of the three monkeys (data not shown).
Recovery of labeled primate or human A. actinomycetemcomitans from oral cavity of primates using the full four-week experimental protocol
As seen in Figure 4, p < 0.05) monkey A. actinomycetemcomitans (Strain Rh 4.35) was recovered at all time points from the 4 of the 5 monkeys and in all 5 monkeys at week 4. Importantly, at no time was there any evidence that labeled A. actinomycetemcomitans derived from humans (IDH 781 SpecR) could be recovered from any collection site. It should be noted that collection at weeks 1 to 3 were done by means of a collection brush that was swiped along the teeth from one quadrant in one case, along the tongue in the second case, and along the cheek in the third collection. Collection at week 4 was done with a scaler and collection was not limited to any tooth surface, hence full mouth plaque collection should have increased the efficiency of the recovery. Nevertheless, despite modest collection procedures performed on weeks 1 to 3, labeled A. actinomycetemcomitans derived from primates was detected in plaque in 4 of the 5 monkeys during that time period. The one Rh monkey where there was a failure to detect labeled A. actinomycetemcomitans did show A. actinomycetemcomitans at the final 4-week collection time. The implanted SpecR A. actinomycetemcomitans colonization level represented anywhere from 1 to 10% of the total A. actinomycetemcomitans flora recovered (data not shown). However, no labeled A. actinomycetemcomitans was collected from primate tongue or cheek at any time-point.
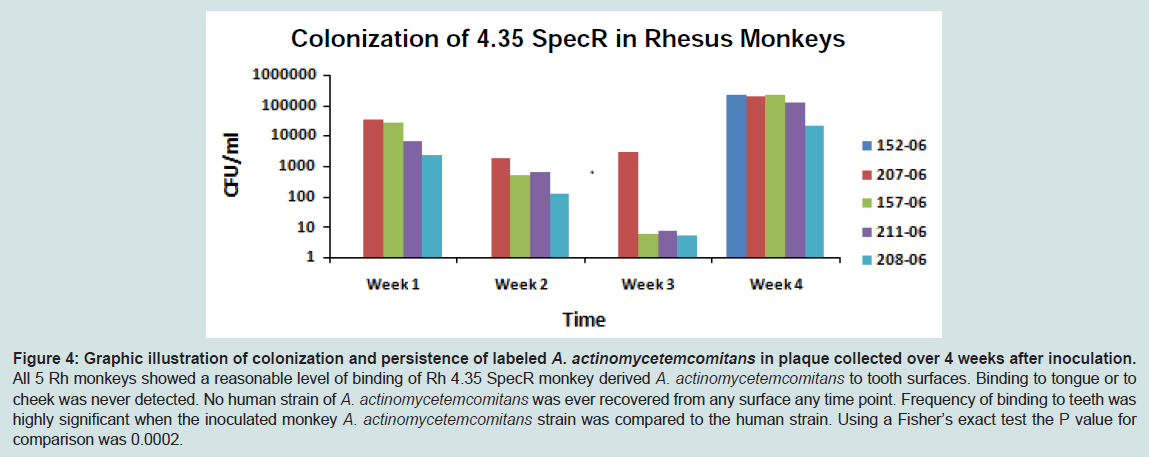
Figure 4: Graphic illustration of colonization and persistence of labeled A. actinomycetemcomitans in plaque collected over 4 weeks after inoculation. All 5 Rh monkeys showed a reasonable level of binding of Rh 4.35 SpecR monkey derived A. actinomycetemcomitans to tooth surfaces. Binding to tongue or to cheek was never detected. No human strain of A. actinomycetemcomitans was ever recovered from any surface any time point. Frequency of binding to teeth was highly significant when the inoculated monkey A. actinomycetemcomitans strain was compared to the human strain. Using a Fisher’s exact test the P value for comparison was 0.0002.
Discussion
We had previously developed a system designed to study A. actinomycetemcomitans in a Specific Pathogen Free rat model where we had created antibiotic labeled A. actinomycetemcomitans strains that were inoculated into the mouths of rats with a reduced oral flora [7]. Over the years we have been able to identify factors required for A. actinomycetemcomitans colonization and persistence in this model [28]. These studies have shown the importance of adherence factors and leukotoxin even in an animal system where A. actinomycetemcomitans is not routinely found [9]. These studies have also indicated that different rat strains show differing sensitivity to the same strain of A. actinomycetemcomitans and that differing strains of A. actinomycetemcomitans produce different levels of disease in the same rat strain [9,28]. Results from these rat studies are very similar to results seen in studies of Porphyromonas gingivalis induced periodontal disease in mice [29,30]. These rodent studies parallel human observational studies showing that not all humans infected with A. actinomycetemcomitans succumb to disease [26,31]. However, since A. actinomycetemcomitans is not typically found in the mouths of Specific Pathogen Free rats it is unlikely that these rodent studies will uncover information related to colonization resistance, the impact other members of the indigenous dental biofilm flora have on A. actinomycetemcomitans growth and survival, or how leukotoxin protects A. actinomycetemcomitans from PMNs, macrophages and lymphocytes [4,32].Recently Hogland Aberg et al. have shown that A. actinomycetemcomitans leukotoxicity substantially enhances the Odds Ratio for onset and progression of LAP in adolescents colonized with A.actinomyetemcomitans [33]. The relationship between leukotoxin and onset and progression of LAP was originally shown by Haubek et al. and this positive correlation between A. actinomycetemcomitans leukotoxic strains and aggressive periodontal disease has been well established over the last several years [31,34]. It is unlikely that the response to A. actinomycetemcomitans leukotoxin in rats can be compared to lymphocytes derived from primates and/or humans [4]. Since the specificity of leukotoxic activity is similar in primates and humans, the primate model is ideal for studying A.actinomycetemcomitans and its relationship to disease. Further, the primate model can help address the question as to how moderate to low leukotoxin producing strains of A. actinomycetemcomitans, are related to aggressive periodontal disease in subjects who do not harbor the high leukotoxin producing strains of A. actinomycetemcomitans [26]. In this case it is likely that a key element in disease progression relates to other members of the flora that either support or interfere with A. actinomycetemcomitans colonization and persistence [26]. This is also best accomplished in an environment that closely resembles that seen in the human mouth. In contrast to what we see in Specific Pathogen Free rats, A. actinomycetemcomitans is a natural inhabitant of the oral flora of the Macaca mulatta (Rhesus macaques).
Our assumptions related to A. actinomycetemcomitans in the mouths of primates is supported by early work by Taichman and colleagues, Beighton et al. and Kilian and Rolla and a number of studies by Ebersole and associates all indicating that A. actinomycetemcomitans is routinely present in the mouths of old world primates [11,12,17,18]. As such, implanting a labeled strain of A. actinomycetemcomitans into the mouth of an animal that supports its growth, survival and response to key virulence factors (i.e. leukotoxin) overtime should allow us to study; 1) factors that support A. actinomycetemcomitans growth and colonization, and perhaps more importantly 2) factors that restrict A. actinomycetemcomitans growth and colonization. Thus the first goal, in the first of several studies, was to establish a reliable model system that would allow us to track A. actinomycetemcomitans overtime in an oral cavity with anatomic and environmental similarities to that found in man [12].
To accomplish this goal we had to develop the optimal conditions for delivery of a “foreign and labeled” A. actinomycetemcomitans strain into the oral cavity of a group of Rh monkeys that were individually housed. It was our contention that selection of the appropriate Rh strain for comparison to our well studied human strain IDH 781 could provide us with information concerning specific A. actinomycetemcomitans virulence traits that support or inhibit its growth and survival in a competitive environment [21,35].
Our first effort was to determine general genomic and functional similarities or differences in key adherence properties in human and primate A. actinomycetemcomitans strains chosen as potential candidate strains for implantation into the mouths Rh monkeys [27]. Rh strain 4.35 was selected by virtue of its attachment characteristics. After choosing this strain we sought to conduct studies aimed at demonstrating reproducible ways of removing or significantly reducing members of the resident flora from tooth and tissue surfaces that might impede colonization of our implanted strain. By reducing the indigenous flora we envisioned that we would provide our implanted strain with a survival advantage. These methods proved to be less straightforward than anticipated. Only methods that included intense scaling and prophylaxis followed by application of topical antiseptics allowed for sufficient microbial reduction to permit colonization of the newly implanted A. actinomycetemcomitans strains. These difficulties were not totally unexpected since the clinical literature is replete with indications regarding the difficulty clinicians have in removal and elimination of A. actinomycetemcomitans from the oral cavity of those infected with A. actinomycetemcomitans[1,36-38]. Nevertheless, the intensity of the recolonization of native A. actinomycetemcomitans was surprising in light of the massive inoculation procedure we used to implant the “foreign” A. actinomycetemcomitans.
Most interesting is the fact that the human strains of A. actinomycetemcomitans inoculated into the mouths of the monkeys were not seen at any time over the 4 week period while all 5 monkeys inoculated with the monkey strain of A. actinomycetemcomitans were colonized. These colonies persisted at reasonable levels throughout the 4-week period. It should be mentioned that feeding alone was not sufficient but topical inoculation of a very high dose of A. actinomycetemcomitans immediately after scaling, prophylaxis and antiseptic application followed by repeated pancake feeding for 5 days did suffice. Also it should be noted that A. actinomycetemcomitans was consistently recovered from tooth surfaces but never from the tongue or cheek surfaces. It is possible that the primate strain chosen possessed superior attachment characteristics when compared to the human strain chosen, although this same human strain had outstanding binding properties in vitro and was successful in its ability to colonize rats in previous experiments [28]. These results suggest a high level of species and tissue or ecological specificity. Exactly why the human and monkey strains of A. actinomycetemcomitans performed so differently in vivo in the implantation experiments is still to be determined. It is also interesting to note that the implanted and successful colonizing strain (Rh 4.35) only formed a low percentage of the total A. actinomycetemcomitans found in monkey plaque. This is especially interesting because of the high dose and repeated inoculation protocol followed in these experiments.
These results suggest that precise discriminatory events take place in the in vivo environment that protects mammals from invasion of alien species (in this case a human strain of A. actinomycetemcomitans) into a biofilm community. This discrimination occurs even when the implanted species shows a high level of similarity to the native strains and even though the indigenous microbial community has been severely compromised by physical and chemical means [4,39].
In conclusion, we feel that we have successfully developed an animal model that has many similarities to the human condition and thus can be used for implantation and study of factors that can influence A. actinomycetemcomitans colonization. The data from this study indicates that intensive physical and chemical cleaning needs to be performed prior to implantation. The data also suggests that implantation is best accomplished by high dose local inoculation of freshly grown strains followed by a feeding protocol that continues the inoculation regimen. Further, this study clearly demonstrates that a monkey strain as opposed to a human strain has superior success in colonization and persistence in the oral cavity of a primate host. With these results in hand we feel that we are positioned to study the deletion of specific A. actinomycetemcomitans virulence genes as well as the effect of commensal bacteria on colonization and survival in the context of a fully developed but reduced oral flora.
Acknowledgements
The authors would like to acknowledge support derived from grant R21 DE 021172 from the National Institutes of Health, National Institute for Craniofacial and Dental Research for support for the work performed in this study. In addition the help provided by members of the staff at the New England Primate Research Center that includes; Ernest Neal, Leah Makaron and Keith Mansfield. In addition DHF would like to thank Dean Cecile Feldman Rutgers SDM for added support for these studies.References
- Zambon JJ (1985) Actinobacillus actinomycetemcomitans in human periodontal disease. J Clin Periodontol 12: 1-20.
- Henderson B, Ward JM, Ready D (2010) Aggregatibacter (Actinobacillus) actinomycetemcomitans : a triple A* periodontopathogen? Periodontol 2000 54: 78-105.
- Fine DH, Kaplan JB, Kachlany SC, Schreiner HC (2006) How we got attached to Actinobacillus actinomycetemcomitans : A model for infectious diseases. Periodontol 2000 42: 114-157.
- Johansson A (2011) Aggregatibacter actinomycetemcomitans leukotoxin: a powerful tool with capacity to cause imbalance in the host inflammatory response. Toxins 3: 242-259.
- Fine DH, Markowitz K, Fairlie K, Tischio-Bereski D, Ferrandiz J, et al. (2014) Macrophage inflammatory protein-1alpha shows predictive value as a risk marker for subjects and sites vulnerable to bone loss in a longitudinal model of aggressive periodontitis. PLoS One 9: e98541.
- Nobbs AH, Jenkinson HF, Jakubovics NS (2011) Stick to your gums: mechanisms of oral microbial adherence. J Dent Res 90: 1271-1278.
- Fine DH, Goncharoff P, Schreiner H, Chang KM, Furgang D, et al. (2001) Colonization and persistence of rough and smooth colony variants of Actinobacillus actinomycetemcomitans in the mouths of rats. Arch Oral Biol 46: 1065-1078.
- Fine DH, Furgang D, McKiernan M, Rubin M (2013) Can salivary activity predict periodontal breakdown in A. actinomycetemcomitans infected adolescents? Arch Oral Biol 58: 611-620.
- Schreiner H, Li Y, Cline J, Tsiagbe VK, Fine DH (2013) A comparison of Aggregatibacter actinomycetemcomitans (Aa) virulence traits in a rat model for periodontal disease. PLoS One 8: e69382.
- Schreiner HC, Sinatra K, Kaplan JB, Furgang D, Kachlany SC, et al. (2003) Tight-adherence genes of Actinobacillus actinomycetemcomitans are required for virulence in a rat model. Proc Natl Acad Sci U S A 100: 7295-7300.
- Ebersole JL, Holt SC, Delaney JE (2014) Acquisition of oral microbes and associated systemic responses of newborn nonhuman primates. Clin Vaccine Immunol 21: 21-28.
- Taichman NS, Simpson DL, Sakurada S, Cranfield M, DiRienzo J, et al. (1987) Comparative studies on the biology of Actinobacillus actinomycetemcomitans leukotoxin in primates. Oral Microbiol Immunol 2: 97-104.
- Fine DH, Velliyagounder K, Furgang D, Kaplan JB (2005) The Actinobacillus actinomycetemcomitans autotransporter adhesin Aae exhibits specificity for buccal epithelial cells from humans and old world primates. Infect Immun 73: 1947-1953.
- Hajishengallis G, Darveau RP, Curtis MA (2012) The keystone-pathogen hypothesis. Nat Rev Microbiol 10: 717-725.
- Jakubovics NS, Kolenbrander PE (2010) The road to ruin: the formation of disease-associated oral biofilms. Oral Dis 16: 729-739.
- Fine DH, Markowitz K, Furgang D, Velliyagounder K (2010) Aggregatibacter actinomycetemcomitans as an early colonizer of oral tissues: epithelium as a reservoir? J Clin Microbiol 48: 4464-4473.
- Kilian M, Rolla G (1976) Initial colonization of teeth in monkeys as related to diet. Infect Immun 14: 1022-1027.
- Beighton D, Taichman NS, Simpson DL, Dirienzo JM, Johnson NW (1989) Intra-oral colonization of macaque monkeys by Actinobacillus actinomycetemcomitans. Oral Microbiol Immunol 4: 35-40.
- Ebersole JL, Cappelli D, Holt SC, Singer RE, Filloon T (2000) Gingival crevicular fluid inflammatory mediators and bacteriology of gingivitis in nonhuman primates related to susceptibility to periodontitis. Oral Microbiol Immunol 15: 19-26.
- Karched M, Furgang D, Sawalha N, Fine DH (2012) Rapid identification of oral isolates of Aggregatibacter actinomycetemcomitans obtained from humans and primates by an ultrafast super convection based polymerase chain reaction. J Microbiol Methods 89: 71-75.
- Hajishengallis G, Lamont RJ (2012) Beyond the red complex and into more complexity: the polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol Oral Microbiol 27: 409-419.
- Hajishengallis G (2014) The inflammophilic character of the periodontitis-associated microbiota. Mol Oral Microbiol 29: 248-257.
- Fine DH, Furgang D, Kaplan J, Charlesworth J, Figurski DH (1999) Tenacious adhesion of Actinobacillus actinomycetemcomitans strain CU1000 to salivary-coated hydroxyapatite. Arch Oral Biol 44: 1063-1076.
- Fine DH, Kaplan JB, Furgang D, Karched M, Velliyagounder K, et al. (2010) Mapping the epithelial-cell-binding domain of the Aggregatibacter actinomycetemcomitans autotransporter adhesin Aae. Microbiology 156: 3412-3420.
- Goncharoff P, Figurski DH, Stevens RH, Fine DH (1993) Identification of Actinobacillus actinomycetemcomitans : polymerase chain reaction amplification of lktA-specific sequences. Oral Microbiol Immunol 8: 105-110.
- Fine DH, Markowitz K, Fairlie K, Tischio-Bereski D, Ferrendiz J, et al. (2013) A consortium of Aggregatibacter actinomycetemcomitans, Streptococcus parasanguinis, and Filifactor alocis is present in sites prior to bone loss in a longitudinal study of localized aggressive periodontitis. J Clin Microbiol 51: 2850-2861.
- Karched M, Furgang D, Planet PJ, DeSalle R, Fine DH (2012) Genome sequence of Aggregatibacter actinomycetemcomitans RHAA1, isolated from a rhesus macaque, an Old World primate. J Bacteriol 194: 1275-1276.
- Schreiner H, Markowitz K, Miryalkar M, Moore D, Diehl S, et al. (2011) Aggregatibacter actinomycetemcomitans-induced bone loss and antibody response in three rat strains. J Periodontol 82: 142-150.
- Baker PJ, Evans RT, Roopenian DC (1994) Oral infection with Porphyromonas gingivalis and induced alveolar bone loss in immunocompetent and severe combined immunodeficient mice. Arch Oral Biol 39: 1035-1040.
- Baker PJ, Dixon M, Evans RT, Roopenian DC (2000) Heterogeneity of Porphyromonas gingivalis strains in the induction of alveolar bone loss in mice. Oral Microbiol Immunol 15: 27-32.
- Haubek D, Poulsen K, Kilian M (2007) Microevolution and patterns of dissemination of the JP2 clone of Aggregatibacter (Actinobacillus) actinomycetemcomitans . Infect Immun 75: 3080-3088.
- Kelk P, Abd H, Claesson R, Sandstrom G, Sjostedt A, et al. (2011) Cellular and molecular response of human macrophages exposed to Aggregatibacter actinomycetemcomitans leukotoxin. Cell Death & Disease 2: e126.
- Hoglund Aberg C, Kwamin F, Claesson R, Dahlen G, Johansson A, et al. (2014) Progression of attachment loss is strongly associated with presence of the JP2 genotype of Aggregatibacter actinomycetemcomitans: a prospective cohort study of a young adolescent population. J Clin Periodontol 41: 232-241.
- Haubek D, Ennibi OK, Poulsen K, Vaeth M, Poulsen S, et al. (2008) Risk of aggressive periodontitis in adolescent carriers of the JP2 clone of Aggregatibacter (Actinobacillus) actinomycetemcomitans in Morocco: a prospective longitudinal cohort study. Lancet 371: 237-242.
- Hooper LV, Macpherson AJ (2010) Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat Rev Immunol 10: 159-169.
- Kornman KS, Caffesse RG, Nasjleti CE (1980) The effect of intensive antibacterial therapy on the sulcular environment in monkeys. Part I. Changes in the bacteriology of the gingival sulcus. J Periodontol 51: 34-38.
- Christersson LA, Slots J, Rosling BG, Genco RJ (1985) Microbiological and clinical effects of surgical treatment of localized juvenile periodontitis. J Clin Periodontol 12: 465-476.
- Takamatsu N, Yano K, He T, Umeda M, Ishikawa I (1999) Effect of initial periodontal therapy on the frequency of detecting Bacteroides forsythus, Porphyromonas gingivalis, and Actinobacillus actinomycetemcomitans. J Periodontol 70: 574-580.
- Hooper LV (2009) Do symbiotic bacteria subvert host immunity? Nat Rev Microbiol 7: 367-374.