
Research Article
*Address for Correspondence: Kyeong Mee Park, Inno Oriental Clinic, Gwanpyeong-Dong, Yusung-Gu, Daejon 305-509, South Korea, Tel: +82 42 671 9933; Fax: +82 42 671 9969; E-mail: kmpark02@empas.com
Citation: Byun B, Suk J, Kwak SS, Lim JS, Choi HJ, et al. Inactivation of mTORC1 Pathway by Aqueous Extract of Acanthopanax senticosus. J Integrative Med Ther. 2015;2(1): 5.
Copyright © 2014 Byun B, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Integrative Medicine & Therapy | Volume: 2, Issue: 1
Submission: 02 June 2015 | Accepted: 16 June 2015 | Published: 22 June 2015
Reviewed & Approved by: Dr. Harold H. Fain, Assistant Professor of Community Medicine, University of North Texas Health Science Center, USA
Reagents and antibodies
At the same time, the inhibitory effect of mTOR/S6K activation by the aqueous extract of A. senticosus correlated with the increased phosphorylations of Akt at Thr 308 by 3-phosphoinositidedependent protein kinase and at Ser 473 by mTORC2 (Figure 1C). Therefore, the presence of negative feedback loops between IRS-1/ PI3K and mTORC1 as well as between mTORC2 and mTORC1 [4] was demonstrated in the cells treated with aqueous extract of A. senticosus Rapamycin, a known inhibitor of mTOR pathway, inhibits differentiation of primary human adipocytes and prevents nutrientmediated insulin resistance in skeletal muscle and adipocyte in vitro [15,16]. In addition, rapamycin protects against high fat-induced obesity in C57BL/6J mice and ameliorates age-dependent obesity in mice [17]. However, a long-term administration of rapamycin is found to cause hyperlipidemia, reduction of fat mass, glucose intolerance, and diabetes-like syndrome by its chronic inhibitory effect on mTORC1/S6K signaling [18,19]. As yet, the underlying mechanism is not clearly elucidated for the glucose intolerance induced by the chronic rapamycin administration. However, it is supposed that the upregulation of hepatic gluconeogenesis and impairment of lipid deposition in adipose tissue possibly underlie the hyperlipidemia and glucose-intolerance in the mice administered with rapamycin for 15days [18].
Inductions of autophagy and cell cycle arrest by A. senticosus extract
Inactivation of mTORC1 Pathway by Aqueous Extract of Acanthopanax senticosus
Boohyeong Byun1, Jinkyu Suk2, Sang Su Kwak2, Jong-Soon Lim3, Hue-Jin Choi5, Tae Hwan Cho2 and Kyeong Mee Park2,5*
- 1Department of Oriental Medicine, Daegu Haany University, Daegu 705-060, South Korea
- 2Department of Biological Sciences, Korea Advanced Institute of Science and Technology, Daejeon 305-701, South Korea
- 3Institute of Traditional Medicine and Bioscience, Daejeon University, Daejeon 300-716, South Korea
- 4Department of Neurosurgery, School of Medicine, Dong-A University, Busan 602-715, South Korea
- 5Inno Oriental Clinic, Daejon 305-509, South Korea
*Address for Correspondence: Kyeong Mee Park, Inno Oriental Clinic, Gwanpyeong-Dong, Yusung-Gu, Daejon 305-509, South Korea, Tel: +82 42 671 9933; Fax: +82 42 671 9969; E-mail: kmpark02@empas.com
Citation: Byun B, Suk J, Kwak SS, Lim JS, Choi HJ, et al. Inactivation of mTORC1 Pathway by Aqueous Extract of Acanthopanax senticosus. J Integrative Med Ther. 2015;2(1): 5.
Copyright © 2014 Byun B, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Integrative Medicine & Therapy | Volume: 2, Issue: 1
Submission: 02 June 2015 | Accepted: 16 June 2015 | Published: 22 June 2015
Reviewed & Approved by: Dr. Harold H. Fain, Assistant Professor of Community Medicine, University of North Texas Health Science Center, USA
Abstract
Dysfunction of mammalian target of rapamycin complex 1 (mTORC1), a central regulator of cell growth, is associated with various diseases including obesity and diabetes. Among ˃ 2,800 plant products used for oriental medicine, Acanthopanax senticosus (A. senticosus) was found to inhibit mTORC1 to phosphorylate S6 kinsase (S6K) in HeLa cells as well as the brain, liver and muscle tissues in diabetic db/db mice. A. senticosus-mediated mTORC1 activity was reversible unlike the prolonged suppression of mTORC1 by rapamycin when HeLa cells were grown in fresh media after the removal of the inhibitors. Autophagy, a self-digestion process occurring in response to nutrient limitation, was activated in HeLa GFP-LC3 cells stably expressing GFP-LC3 by the treatment with the aqueous extract of A. senticosus. The conversion of cytoplasmic LCI into lipidated LCII was increased by fivefold in HeLa GFP-LC3 cells treated with A. senticosus extract. Progression of cell cycle was also attenuated at G2/M phase by the treatment of the aqueous extract of A. senticosus. Taken together, these results suggest that the aqueous extract of A. senticosus possiblyprevents obesity-related diabetes by inhibiting mTOR signaling.Keywords
Acanthopanax senticosus; Autophagy; Cell cycle; db/db mouse; mTORC1; S6KIntroduction
The mammalian target of rapamycin (mTOR) is a Ser/Thr kinase that belongs to the phosphatidylinositol kinase-related protein kinase family [1]. mTOR exists in two complexes, rapamycin-sensitive mTORC1 and rapamycin-resistant mTORC2. mTORC1 activates cellular processes by phosphorylating two downstream effectors, ribosome S6 kinases (S6K) and eukaryotic initiation factor 4E-binding proteins (4E-BPs) [2]. Both mTORC1 and S6K are activated by insulin through the insulin receptor/insulin receptor substrate/ phosphatidyinositol 3-kinase/Akt (PI3-kinase/Akt) pathway [3].Chronic activation of mTORC1/S6K pathway by prolonged insulin treatment causes insulin resistance by phosphorylating and degrading insulin receptor substrate 1/2 and feed-back inhibiting the activation of PI3-kinase/Akt pathway [4]. A recent report indicates that mice deficient S6K are protected against diet-induced obesity and show enhanced insulin sensitivity owing to the loss of the negative feedback loop [3].
Acute inhibition of mTORC1 by rapamycin is reportedly known to protect from insulin resistance induced by excessive nutrient [5]. However, a chronic rapamycin administration is paradoxically found to promote glucose intolerance and a diabetes-like syndrome [6]. Moreover, rapamycin has some unwanted side effects such as lymphedema [7], stomatitis [8], and azoospermea [9]. Therefore, the interventions targeting mTOR/S6K signaling without any harmful side effects are sought for the prevention of obesity and insulin resistance.
In search of naturally occurring plant products that can modulate mTOR pathway, a large collection of natural plant products used in folk medicine was screened. Among 2,800 plant products, Acanthopanax senticosus (A. senticosus) was found to inhibit mTORC1/S6K activity in both cultured HeLa cells and obese diabetic db/db mice. At the same time, the aqueous extract of A. senticosus induced autophagy and arrested cell cycle at G2/M phase in cultured HeLa cells. In this report, we launch revealing out the mechanism of A. senticosus on the intervention in mTOR signaling.
Materials and Methods
Mouse monoclonal antibody for S6K and Anti-GFP mouse antibody were from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Mouse antibody against phosphorylated S6K (Thr 389) was from Cell Signaling Technology (Boston, MA, USA). Rapamycin and rabbit antibodies for phosphorylated Akt at Thr 308 and Ser 473 were also from Cell Signaling Technology. Anti-α-tubulin antibody (T5168) was purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Preparation of an aqueous extract from roots of A. senticosus
Dried A. senticosus were obtained from Natural Medicine Research Center of Korea Institute of Bioscience and Biotechnology, Daejeon, South Korea. The plant was identified by a botanist from the Institute of Traditional Medicine and Bioscience, Daejeon University, Daejeon, South Korea. Roots of A. senticosus (600 g) were pulverized, extensively washed with water, and dried at 40 °C in a hot air oven. The plant material was homogenized in 2 L of hot boiling water for 2 h and allowed to stand for 24 h. After filtration, the filtrate was lyophilized. The lyophilized extract (~50 g) was dissolved in DMSO and used to evaluate its effect on mTORC1 signaling in both cultured cells and animal model.
Cell culture and immunoblot assay
HeLa cells were cultured in Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 10% (v/v) fetal bovine serum (Gibco, Grand island, NY, USA) and maintained at 37 °C in 5% (v/v) CO2 under a humidified atmosphere. Sample proteins were analyzed by 12% SDS-PAGE and immunoblotting.
Experimental mice
C57BLKS/J-m+/Lepr db (db/db) mice were purchased from Japan SLC, Inc. Experiments using this mouse strain were compliant with the guideline of Institutional Animal Care and Use Committee (IACUC) at Korea Advanced Institute of Science and Technology (KAIST). Diabetic obese db/db mice were fed a normal diet under ad libitum conditions. A. senticosus extract (100 µg/g body weight/day) was intraperitoneally injected to 15-week-old mice (n=3 female and 3 male mice) for a week. For analysis of mTORC1 activity, mice were sacrificed to prepare tissue samples for immunoblotting.
Induction of autophagy
Autophagy induction after the treatment of A. senticosus extract was evaluated by monitoring the conversion of cytoplasmic LC3I into lipidated LC3II [10] in cultured human HeLa GFP-LC3 cells. GFP-LC3 HeLa cells were established according to Suk et al. [11]. The induction of autophagy by the plant extract was quantified by measuring the percentage of GFP-LC3 positive HeLa GFP-LC3 cells with punctuate dots using fluorescence microscopy.
Cell cycle analysis
Cell cycle was analyzed by flow cytometry. HeLa cells treated with A. senticosus extract were fixed in 1% formaldehyde PBS followed by ice-cold 70% ethanol, and incubated on ice for 30 mins with thorough mixing. Fixed cells were then treated with RNaseI and stained with propidium iodide for fluorescence-activated cell sorter (FACS) analysis of DNA contents [12].
Inhibition of mTORC1 pathway by A. senticosus extract
Results and Discussion
In the present study, the effect of A. senticosus extract that inhibits mTORC1 was investigated in both cultured human cells and diabetic obese db/db mice. The aqueous extract of A. senticosus inhibited the ability of mTORC1 to phosphorylate its substrate protein S6K in HeLa ce1ls (Figure 1). In the diabetic db/db mice in which mTORC1 activity is highly activated [13], A. senticosus extract (100 µg/g body weight/day), administered intraperitoneally for a week, induced a down-regulation of S6K in tissues of brain, liver, and muscle (Figure 1b). As a nutrient-dependent stimulation of the mTOR/S6K pathway can induce insulin resistance in skeletal muscle and liver [3,14], the inhibition of S6K phosphorylation in tissues of muscle and liver in the diabetic db/db mice administered with A. senticosus reveals A. senticosus could be used to treat and/or prevent nutrient-dependent obesity and insulin resistance in vivo.
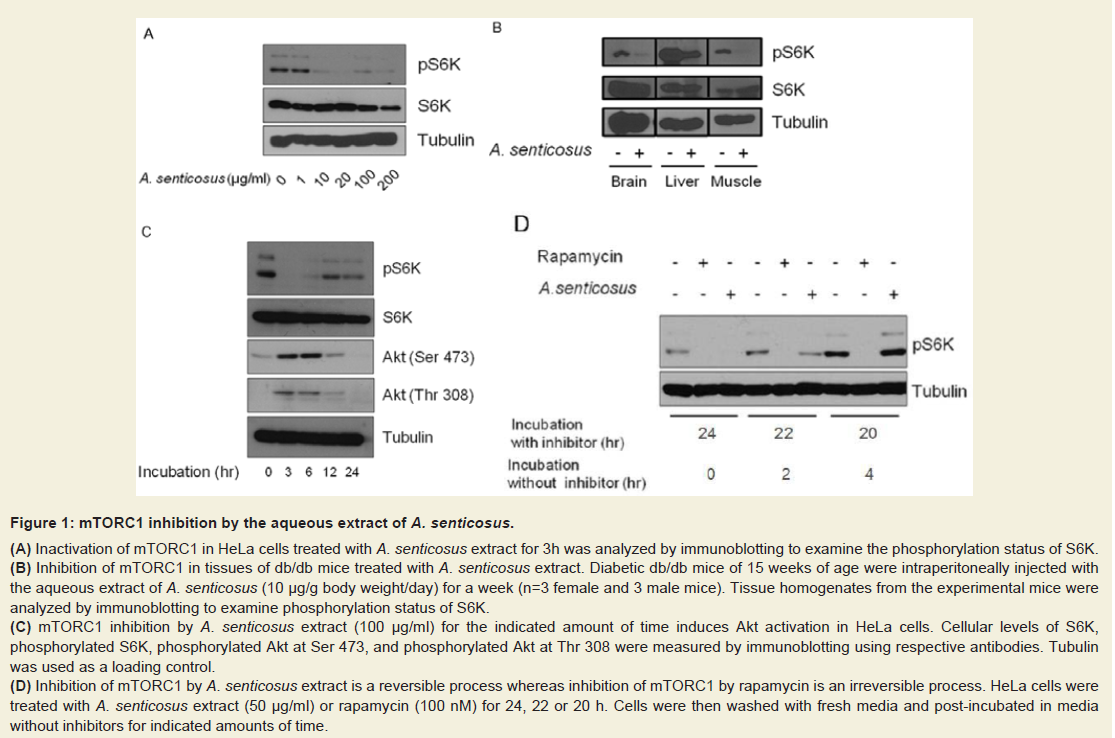
Figure: 1 : mTORC1 inhibition by the aqueous extract of A. senticosus.
(A) Inactivation of mTORC1 in HeLa cells treated with A. senticosus extract for 3h was analyzed by immunoblotting to examine the phosphorylation status of S6K.
(B) Inhibition of mTORC1 in tissues of db/db mice treated with A. senticosus extract. Diabetic db/db mice of 15 weeks of age were intraperitoneally injected withthe aqueous extract of A. senticosus (10 µg/g body weight/day) for a week (n=3 female and 3 male mice). Tissue homogenates from the experimental mice wereanalyzed by immunoblotting to examine phosphorylation status of S6K.
(C) mTORC1 inhibition by A. senticosus extract (100 µg/ml) for the indicated amount of time induces Akt activation in HeLa cells. Cellular levels of S6K,phosphorylated S6K, phosphorylated Akt at Ser 473, and phosphorylated Akt at Thr 308 were measured by immunoblotting using respective antibodies. Tubulinwas used as a loading control.
(D) Inhibition of mTORC1 by A. senticosus extract is a reversible process whereas inhibition of mTORC1 by rapamycin is an irreversible process. HeLa cells weretreated with A. senticosus extract (50 µg/ml) or rapamycin (100 nM) for 24, 22 or 20 h. Cells were then washed with fresh media and post-incubated in mediawithout inhibitors for indicated amounts of time.
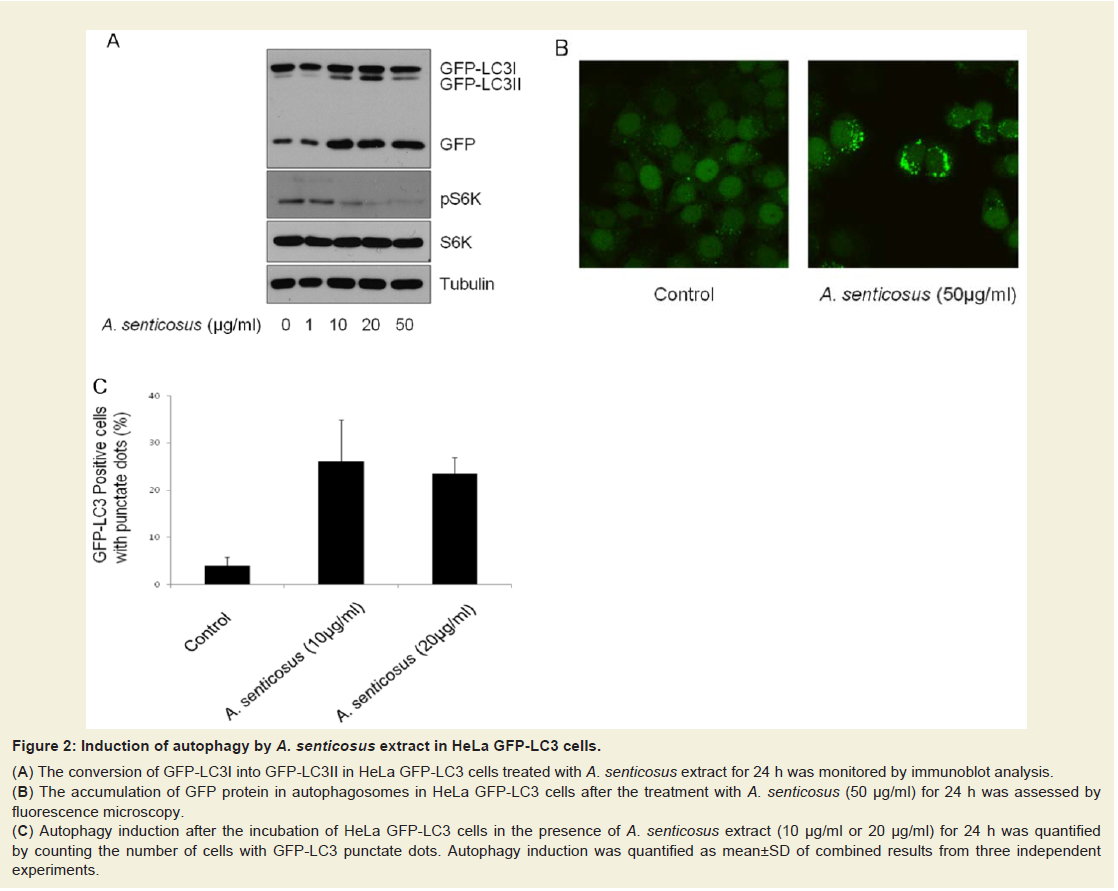
Figure: 2 : Induction of autophagy by A. senticosus extract in HeLa GFP-LC3 cells.
(A) The conversion of GFP-LC3I into GFP-LC3II in HeLa GFP-LC3 cells treated with A. senticosus extract for 24 h was monitored by immunoblot analysis.
(B) The accumulation of GFP protein in autophagosomes in HeLa GFP-LC3 cells after the treatment with A. senticosus (50 µg/ml) for 24 h was assessed byfluorescence microscopy.
(C) Autophagy induction after the incubation of HeLa GFP-LC3 cells in the presence of A. senticosus extract (10 µg/ml or 20 µg/ml) for 24 h was quantifiedby counting the number of cells with GFP-LC3 punctate dots. Autophagy induction was quantified as mean±SD of combined results from three independentexperiments
As HeLa cells were treated for inhibitors for 24, 22, or 20 hours and then incubated in fresh media without inhibitors for 0, 2, or 4 hours respectively, the suppression of mTORC1 activity was persisted only in cells treated with rapamycin but not in the cells treated with A. senticosus (Figure 1d). In the absence of A. senticosus, depressed activity of mTORC1 resumed to normal apart from rapamycin. As revealed in (Figure 1d), if the administration of A. senticosus stops, depressed mTORC1/S6K activity might be resumed thus not to induce hepatic gluconeogenic shunt as observed in rapamycin, which uncouples between hepatic insulin signaling and glucose homeostasis [18].
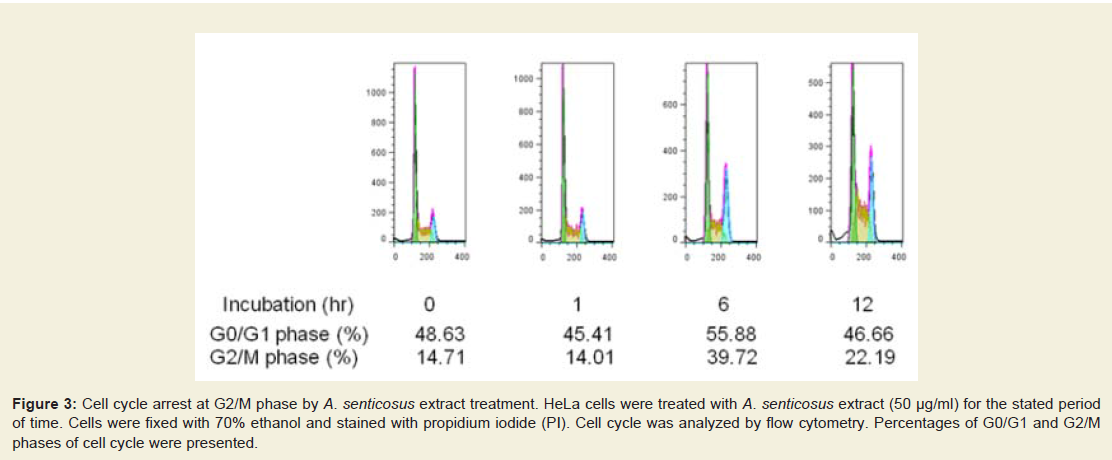
Figure: 3 :Cell cycle arrest at G2/M phase by A. senticosus extract treatment. HeLa cells were treated with A. senticosus extract (50 µg/ml) for the stated periodof time. Cells were fixed with 70% ethanol and stained with propidium iodide (PI). Cell cycle was analyzed by flow cytometry. Percentages of G0/G1 and G2/Mphases of cell cycle were presented.
Autophagy is considered as a protection process which enables cells to survive adverse conditions like nutrient limitation, infections, toxins, and other types of cell stress. Starvation activated liver autophagy, whereas nutrient supplementation following food deprivation suppressed it [20]. Exogenous substrate availability through insulin signals an anabolic state and activates mTORC1 to stimulate protein synthesis and aerobic ATP production. On the other hand, shortage of cellular substrate or dysfunctional oxidative phosphorylation inhibits mTORC1, activating autophagic pathway [20]. In this study, autophagy induction was assessed and quantified in established HeLa GFP-LC3 cells. In response to the treatment of the aqueous extract of A. senticosus to HeLa GFP-LC3 cells, LC3I was converted into LC3II, a known hallmark of autophagy [21], in HeLa GFP-LC3 cells (Figure 2a). The accumulation of GFP-LC3 in the autophagosomes in HeLa GFP-LC3 cells stably expressing LC3 was imaged by fluorescent microscopy (Figure 2B) and quantified (Figure 2c). Treatment of HeLa GFP-LC3 cells with 10 µg/ml aqueous extract of A. senticosus for 24 h increased the number of cells with GFP-LC3 punctate dots by approximately fivefold.
Because autophagy is a catabolic pathway that degrades bulkcytosol in the lysosomal compartments enabling amino acids and fatty acids to be recycled, attenuation in the progression of cell cycle is very plausible. Consistent with the induction of autophagy, the cells treated with A. senticosus failed to progress in normal cell cycle (Figure 3). Whereas rapamycin inhibits growth factor signaling and induces G1 arrest in many cell types [22], A. senticosus extract induced G2 arrest in HeLa cells (Figure 3). As cellular processes like cell growth are directly linked with mTORC1 function that integrates signals for protein synthesis, it is evident that the inhibition of mTORC1 signaling pathway by A. senticosus extract should be functionally linked with the attenuation of cell cycling.
Although our findings add novel information to explore rapamycin analogues in treating obesity and obesity-related diabetes, the exact mechanism of A. senticosus to inhibit mTOR signaling remains to be identified. As A. senticosus extract was chronically administered to diabetic db/db mice fed with normal diet in this experiment, there was no difference in blood glucose concentrations between A. senticosus-fed db/db mice and control db/db mice. We anticipate there might be some remarkable differences between A. senticosus–administered and control db/db mice if the db/db mice are fed with high nutrient diet. Therefore, our next study will focus on the effect of A. senticosus on glucose and lipid metabolisms in high nutrient-fed diabetic db/db mice.
References
- Wullschleger S, Loewith R, Hall MN (2006) TOR signaling in growth and metabolism. Cell 124: 471-484.
- Burnett PE, Barrow RK, Cohen NA, Snyder SH, Sabatini DM (1998) RAFT1 phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc Natl Acad Sci U S A 95: 1432-1437.
- Khamzina L, Veilleux A, Bergeron S, Marette A (2005) Increased activation of the mammalian target of rapamycin pathway in liver and skeletal muscle of obese rats: possible involvement in obesity-linked insulin resistance. Endocrinology 146: 1473-1481.
- Smith LK, Vlahos CJ, Reddy KK, Falck JR, Garner CW (1995) Wortmannin and LY294002 inhibit the insulin-induced down-regulation of IRS-1 in 3T3-L1 adipocytes. Mol Cell Endocrinol 113: 73-81.
- Tzatsos A, Kandror KV (2006) Nutrients suppress phosphatidylinositol 3-kinase/Akt signaling via Raptor-dependent mTOR-mediated insulin receptor substrate 1 phosphorylation. Mol Cell Biol 26: 63-76.
- Cruzado JM (2008) Nonimmunosuppressive effects of mammalian target of rapamycin inhibitors. Transplant Rev (Orlando) 22: 73-81.
- Hille U, Soergel P, Makowski L, Dork-Bousset T, Hillemanns P (2012) Lymphedema of the breast as a symptom of internal diseases or side effect of mTOR inhibitors. Lymphat Res Biol 10: 63-73.
- Pilotte AP, Hohos MB, Polson KM, Huftalen TM, Treister N (2011) Managing stomatitis in patients treated with mammalian target of rapamycin inhibitors. Clin J Oncol Nurs 15: E83-E89.
- Skrzypek J, Krause W (2007) Azoospermia in a renal transplant recipient during sirolimus (rapamycin) treatment. Andrologia 39: 198-199.
- Mizushima N, Klionsky DJ (2007) Protein turnover via autophagy: implications for metabolism. Annu Rev Nutr 27: 19-40.
- Suk J, Kwak SS, Lee JH, Choi JH, Lee SH, et al. (2011) Alkaline stress-induced autophagy is mediated by mTORC1 inactivation. J Cell Biochem 112: 2566-2573.
- Ramirez-Valle F, Badura ML, Braunstein S, Narasimhan M, Schneider RJ (2010) Mitotic raptor promotes mTORC1 activity, G(2)/M cell cycle progression, and internal ribosome entry site-mediated mRNA translation. Mol Cell Biol 30: 3151-3164.
- Mori H, Inoki K, Masutani K, Wakabayashi Y, Komai K, et al. (2009) The mTOR pathway is highly activated in diabetic nephropathy and rapamycin has a strong therapeutic potential. Biochem Biophys Res Commun 384: 471-475.
- Tremblay F, Krebs M, Dombrowsky L, Brehm A, Bernroider E, et al. (2005) Overactivation of S6 kinase 1 as a cause of human insulin resistance during increased amino acid availability. Diabetes 54: 2674-2684.
- Bell A, Grunder L, Sorisky A (2000) Rapamycin inhibits human adipocyte differentiation in primary culture. Obes Res 8: 249-254.
- Chang GR, Chiu YS, Wu YY, Chen WY, Liao JW, et al. (2009) Rapamycin protects against high fat-induced obesity in C57BL/6 mice. J Pharmacol Sci 109: 496-503.
- Yang SB, Tien AC, Boddupalli G, Xu AW, Jan YN, et al. (2012) Rapamycin ameliorates age-dependent obesity associated with increased mTOR signaling in hypothalamic POMC neurons. Neuron 75: 425-436.
- Houde VP, Brûlé S, Festuccia WT, Blanchard PG, Bellmann K, et al. (2010) Chronic rapamycin treatment causes glucose intolerance and hyperlipidemia by upregulating hepatic gluconeogenesis and impairing lipid deposition in adipose tissue. Diabetes 59: 1338-1348.
- Hu X, Sato J, Bajotto G, Khookhor O, Ohsawa I, et al. (2010) Goshajinkigan (Chinese herbal medicine Niu-Che-Sen-Qi-Wan) improves insulin resistance in diabetic rats via the nitric oxide pathway. Nagoya J Med Sci 72: 35-42.
- Seldin MM, Lei X, Tan SY, Stanson KP, Wei Z, et al. (2013) Skeletal muscle-derived myonectin activates the mammalian target of rapamycin (mTOR) pathway to suppress autophagy in liver. J Biol Chem 288: 36073-36082.
- Levine B, Kroemer G (2008) Autophagy in the pathogenesis of disease. Cell 132: 27-42.
- Metcalfe SM, Canman CE, Milner J, Morris RE, Goldman S, et al. (1997) Rapamycin and p53 act on different pathways to induce G1 arrest in mammalian cells. Oncogene 15: 1635-1642.