
Journal of Cancer Sciences
Download PDF
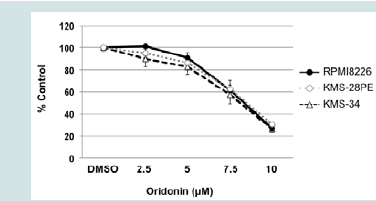 Figure 1A: Oridonin-mediated growth inhibition and cell death in MM cells. KMS-28PE, KMS-34, and RPMI8226 cells were cultured in 96-well tissue culture plates in the presence or absence of oridonin at the indicated concentrations for 24 h. Cell viability was measured using the MTT assay. Data are expressed as the mean percentage of the control cells and represent the average of triplicate samples. The results shown are representative of three independent experiments.
Figure 1A: Oridonin-mediated growth inhibition and cell death in MM cells. KMS-28PE, KMS-34, and RPMI8226 cells were cultured in 96-well tissue culture plates in the presence or absence of oridonin at the indicated concentrations for 24 h. Cell viability was measured using the MTT assay. Data are expressed as the mean percentage of the control cells and represent the average of triplicate samples. The results shown are representative of three independent experiments. 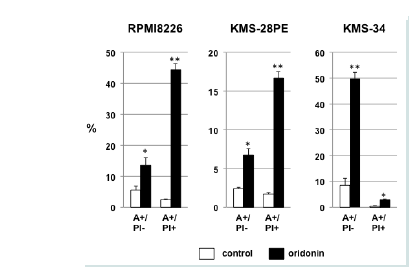 Figure 1B: Oridonin-mediated growth inhibition and cell death in MM cells. KMS-28PE, KMS-34, and RPMI8226 cells were cultured in 6-well tissue culture plates in the presence or absence of oridonin (10 μM) for 24 h. The frequency of cell death was measured using flow cytometry analysis of annexin V/propidium iodide (PI)-stained cells. Data represent the average of triplicate samples, and error bars represent one standard deviation (SD) from the mean. The results shown are representative of two independent experiments.*P<0.05, **P<0.01: oridonin vs. control. A: Annexin V; PI: Propidium Iodide.
Figure 1B: Oridonin-mediated growth inhibition and cell death in MM cells. KMS-28PE, KMS-34, and RPMI8226 cells were cultured in 6-well tissue culture plates in the presence or absence of oridonin (10 μM) for 24 h. The frequency of cell death was measured using flow cytometry analysis of annexin V/propidium iodide (PI)-stained cells. Data represent the average of triplicate samples, and error bars represent one standard deviation (SD) from the mean. The results shown are representative of two independent experiments.*P<0.05, **P<0.01: oridonin vs. control. A: Annexin V; PI: Propidium Iodide. 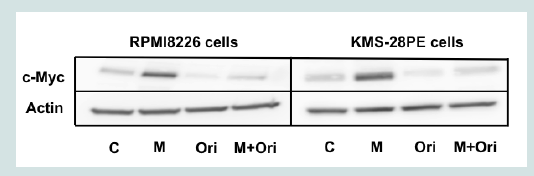 Figure 2A: Oridonin suppresses c-Myc expression and activates caspase-3 and PARP but not IRF-4 and Fbxw7α in MM cells. RPMI8226 and KMS-28PE cells were treated with oridonin (10 μM) for the indicated time (0,12,18, or 24 h) and analyzed using immunoblotting assays with specific antibodies against c-Myc, Mcl-1, procaspase-9, cleaved caspase-3, cleaved PARP, Fbxw7α, and β-actin.
Figure 2A: Oridonin suppresses c-Myc expression and activates caspase-3 and PARP but not IRF-4 and Fbxw7α in MM cells. RPMI8226 and KMS-28PE cells were treated with oridonin (10 μM) for the indicated time (0,12,18, or 24 h) and analyzed using immunoblotting assays with specific antibodies against c-Myc, Mcl-1, procaspase-9, cleaved caspase-3, cleaved PARP, Fbxw7α, and β-actin. 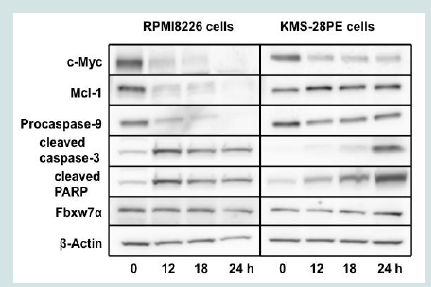 Figure 2B: Oridonin suppresses c-Myc expression and activates caspase-3 and PARP but not IRF-4 and Fbxw7α in MM cells. RPMI8226 and KMS-28PE cells were treated with oridonin (10 μM) for the indicated time (0,2,4, or 6 h) and analyzed using immunoblotting assays with specific antibodies against c-Myc, IRF-4, Mcl-1, Fbxw7α, and β-actin. The immunoblot shown is representative of two independent experiments.
Figure 2B: Oridonin suppresses c-Myc expression and activates caspase-3 and PARP but not IRF-4 and Fbxw7α in MM cells. RPMI8226 and KMS-28PE cells were treated with oridonin (10 μM) for the indicated time (0,2,4, or 6 h) and analyzed using immunoblotting assays with specific antibodies against c-Myc, IRF-4, Mcl-1, Fbxw7α, and β-actin. The immunoblot shown is representative of two independent experiments. 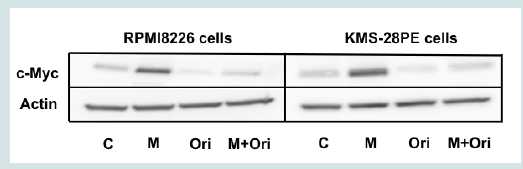 Figure 3A: Oridonin induces the downregulation of c-myc mRNA and the proteasomal degradation of c-Myc in MM cells. MG132 blocked oridonin-mediated suppression of c-Myc levels. RPMI8226 and KMS-28PE cells were incubated in the presence or absence of oridonin (10 μM), MG-132 (10 μM), or both agents for 2 h and analyzed using immunoblotting assays with specific antibodies againstc-Myc and β-actin. The immunoblot shown is representative of two independent experiments. C: Control; M: MG132; Ori: Oridonin.
Figure 3A: Oridonin induces the downregulation of c-myc mRNA and the proteasomal degradation of c-Myc in MM cells. MG132 blocked oridonin-mediated suppression of c-Myc levels. RPMI8226 and KMS-28PE cells were incubated in the presence or absence of oridonin (10 μM), MG-132 (10 μM), or both agents for 2 h and analyzed using immunoblotting assays with specific antibodies againstc-Myc and β-actin. The immunoblot shown is representative of two independent experiments. C: Control; M: MG132; Ori: Oridonin. 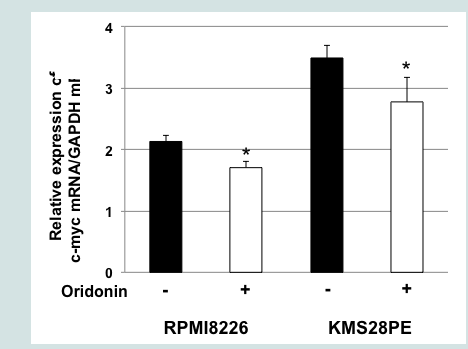 Figure 3B: Oridonin induces the downregulation of c-myc mRNA and the proteasomal degradation of c-Myc in MM cells. c-myc mRNA levels decreased in oridonin-treated MM cells. RPMI8226 and KMS-28PE cells were treated with oridonin (10 μM) for 2 h, and total RNA was subsequently prepared. Relative expression levels of c-myc mRNA were quantified using real-time RT-PCR analysis. The data represent the average of triplicate samples, and error bars represent one SD from the mean. The results shown are representative of two independent experiments. *P<0.05: oridonin vs. control.
Figure 3B: Oridonin induces the downregulation of c-myc mRNA and the proteasomal degradation of c-Myc in MM cells. c-myc mRNA levels decreased in oridonin-treated MM cells. RPMI8226 and KMS-28PE cells were treated with oridonin (10 μM) for 2 h, and total RNA was subsequently prepared. Relative expression levels of c-myc mRNA were quantified using real-time RT-PCR analysis. The data represent the average of triplicate samples, and error bars represent one SD from the mean. The results shown are representative of two independent experiments. *P<0.05: oridonin vs. control. 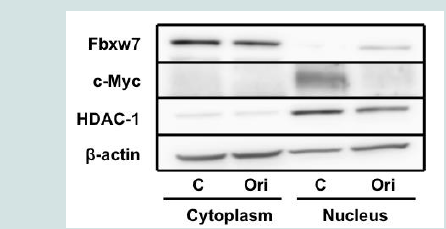 Figure 4A: Oridonin induces the nuclear accumulation of Fbxw7α in RPMI8226 cells. (A). RPMI8226 cells were treated with oridonin (10 μM) for 6 h and analyzed using immunoblotting assays with specific antibodies against Fbxw7α, c-Myc, HDAC1, and β-actin. The immunoblot shown is representative of two independent experiments. C: Control; Ori: Oridonin.
Figure 4A: Oridonin induces the nuclear accumulation of Fbxw7α in RPMI8226 cells. (A). RPMI8226 cells were treated with oridonin (10 μM) for 6 h and analyzed using immunoblotting assays with specific antibodies against Fbxw7α, c-Myc, HDAC1, and β-actin. The immunoblot shown is representative of two independent experiments. C: Control; Ori: Oridonin. 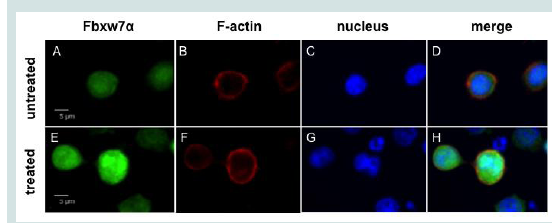 Figure 4B: Oridonin induces the nuclear accumulation of Fbxw7α in RPMI8226 cells. (Top (A-D) and bottom (E-H) panels show untreated and oridonin-treated RPMI8226 cells, respectively. Images were acquired using fluorescence microscopy. Fbxw7α is shown in green (A,E), F-actin in red (B,F), and the nucleus in blue (C,G). Merged images are shown in panels D and H.
Figure 4B: Oridonin induces the nuclear accumulation of Fbxw7α in RPMI8226 cells. (Top (A-D) and bottom (E-H) panels show untreated and oridonin-treated RPMI8226 cells, respectively. Images were acquired using fluorescence microscopy. Fbxw7α is shown in green (A,E), F-actin in red (B,F), and the nucleus in blue (C,G). Merged images are shown in panels D and H.
Research Article
Oridonin Induces Apoptosis of Myeloma Cells by Inducing C-Myc Degradation via Nuclear Accumulation of Fbxw7α
Motoki Takano1, Maki Nishiya1, Shugo Kowata1, Yoji Ishida1 and Shigeki Ito2*
- 1Department of Internal Medicine, Iwate Medical University School of Medicine, Japan
- 2Department of Clinical Oncology, Iwate Medical University School of Medicine, Japan
*Address for Correspondence: Shigeki Ito, Department of Clinical Oncology, Iwate Medical University School of Medicine, 19-1 Uchimaru, Morioka, Iwate 020-8505, Japan, Tel: +81-19-651-5111 (Ext.3379); Fax: +81-19-651-5185; E-mail: shigei@iwate-med.ac.jp
Citation: Takano M, Nishiya M, Kowata S, Ishida Y, Ito S. Oridonin Induces Apoptosis of Myeloma Cells by Inducing C-Myc Degradation via Nuclear Accumulation of Fbxw7α. J Cancer Sci. 2017;4(1): 6.
Journal of Cancer Sciences | ISSN: 2377-9292 | Volume: 4, Issue: 1
Submission: 28 October 2017| Accepted: 29 November 2017 | Published: 05 December 2017
Copyright: © 2017 Takano M, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
The overall survival of patients with multiple myeloma has recently improved, primarily due to the use of several novel agents, including proteasome inhibitors and immunomodulatory drugs. However, as MM remains incurable due to its eventual relapse, the development of more effective agents is greatly needed. Oridonin is a diterpenoid that induces apoptosis and autophagy in various types of cancer. However, the effects of oridonin on MM cells have not been fully elucidated. In the present study, we found that oridonin inhibited proliferation and induced caspase-dependent apoptosis in MM cells. Interestingly, oridonin suppressed c-Myc protein levels and promoted the nuclear accumulation of F-box and WD-40 domain protein 7α, a component of the SCF-like ubiquitin ligase complex, suggesting that oridonin induced c-Myc down regulation might at least in part be due to proteasomal degradation of c-Myc via Fbxw7α-mediated ubiquitination. These results suggest that the effects of oridonin in MM patients in the clinical setting warrant investigation.
Keywords
c-myc; Multiple myeloma; Fbxw7α; Oridonin
Abbreviations
Abbreviations: MM: Multiple Myeloma; Fbxw7: F-box and WD-40 domain protein 7; SCF: Skp, Cullin, and F-box; DMSO: Dissolved in Dimethyl Sulfoxide; PARP: Poly ADP-Ribose Polymerase; IRF: Interferon Regulatory Factor; HDAC1: Histone Deacetylase 1; FBS: Fetal Bovine Serum; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide; PI: Propidium Iodide; RT-PCR: Reverse Transcription Polymerase Chain Reaction; PBS: Phosphate-buffered Saline; SD: Standard Deviation
Introduction
Multiple Myeloma (MM) is a B-cell malignancy characterized by the accumulation of neoplastic plasma cells that typically secrete monoclonal proteins. The overall survival of patients with MM has significantly improved over the last decade, primarily due to the use f novel agents such as thalidomide, lenalidomide and bortezomib, as well as the combined use of high-dose chemotherapy with autologous hematopoietic stem cell transplantation [1,2]. However, MM remains incurable, as patients frequently relapse even after a complete response is achieved [1,2]. Therefore, the development of newer agents with unique mechanisms of action is needed to further improve outcomes in MM patients.
c-Myc is a helix-loop-helix leucine zipper transcription factor that plays essential roles in cell growth, proliferation, survival, and metabolic adaptation [3]. c-Myc is deregulated in multiple types of cancer, including leukemia and lymphoma [4]. In addition, c-Myc is activated in approximately two-thirds of patients with MM but not in patients with monoclonal gammopathy of undetermined significance, indicating that c-Myc plays a pivotal role in myeloma pathogenesis [5]. A recent study demonstrated that MM cells depend on c-Myc for survival, as the ablation of c-myc using small hairpin RNAs or small molecule inhibitors induced MM cell death [6]. We previously demonstrated that the survivin inhibitor YM155 suppressed proliferation and survival in MM cells by promoting the proteasomal degradation of c-Myc [7]. Thus, c-Myc is an attractive target for myeloma treatment.
Oridonin is a natural diterpenoid isolated from the Chinese medicinal herb Rabdosia rubescens. Oridonin has been shown to induce apoptosis and autophagy in a wide variety of cancer cells, including those isolated from prostate, breast, lung, and lymphoid malignancies [8-11]. However, the mechanisms underlying the effects of oridonin in MM cells remain largely unknown. Huang et al. found that oridonin induced apoptosis in human erythroleukemia K562 cells by inducing the proteasomal degradation of c-Myc by F-box and WD-40 domain protein 7 (Fbxw7), a component of the Skp, Cullin, and F-box (SCF)-like ubiquitin ligase complex [10]. Zeng et al. demonstrated that oridonin induced apoptosis and autophagy in human MM RPMI8226 cells by regulating the generation of intracellular reactive oxygen species generation and sirtuin 1 [11]. In the present study, we demonstrated that oridonin suppresses cell proliferation and induces caspase-dependent apoptosis in MM cells. Oridonin-mediated these effects, at least in part, by inducing the nuclear translocation of Fbxw7α, thereby promoting Fbxw7- mediated proteasomal degradation of c-Myc.
Materials and Methods
Reagents and antibodies
Oridonin was purchased from Sigma-Aldrich (St. Louis, MO), and the proteasome inhibitor MG132 was purchased from EMD Biosciences (La Jolla, CA). Both agents were dissolved in Dimethyl Sulfoxide (DMSO), and working solutions were prepared by diluting the stock solution in RPMI 1640 medium. Control cells were treated with 0.1% DMSO in complete medium, as this was the maximum final DMSO concentration in oridonin-treated cells. Antibodies against Mcl-1, c-Myc, cleaved caspase-3, procaspase-9, cleaved Poly ADPribose Polymerase (PARP), Interferon Regulatory Factor (IRF)-4, and β-actin were obtained from cell signaling technology (Beverly, MA). Antibodies against Fbxw7α were purchased from Allele Biotechnology (San Diego, CA). Antibodies against Histone Deacetylase 1 (HDAC1) were purchased from Santa Cruz Biotechnology (Dallas, TX).
Cell lines
The human myeloma cell line RPMI8226 (American Type Culture Collection [ATCC], Rockville, MD) was derived from the peripheral blood of a 61-year-old male with MM, and reveals hyper diploid karyotype and has no high-risk cytogenetic features [12]. The myeloma cell lines KMS-28PE and KMS-34 were kindly provided by Dr. Tamura (Nippon Medical School, Tokyo, Japan). KMS-28PE and KMS-34 cells were derived from pleural effusions from a 77-yearfemale and a 60-year-female with t(4;14) myeloma, respectively, and were established at Kawasaki Medical School, Okayama, Japan [13,14]. MM cell lines were cultured in RPMI 1640 (Gibco, Grand Island, NY) supplemented with 10% Fetal Bovine Serum (FBS) (HyClone, Logan, UT), 100 U/ml penicillin (Gibco), 100 μg/ml streptomycin (Gibco), and 2 mM glutamine (Gibco) in a 5% CO 2 incubator at 37°C.
Cell proliferation assay
Cell proliferation assays were conducted as previously described [15]. Briefly, 2×105 cells/ml from each line were cultured in RPMI 1640 medium supplemented with 10% FBS in the presence or absence of the indicated concentrations of oridonin for 24 h. The 3-(4,5-dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) assay was subsequently conducted according to the manufacturer’s recommendations.
Apoptosis assay
The apoptosis assay was conducted as previously described[14]. Briefly, RPMI8226, KMS-28PE, and KMS-34 cells were plated at a concentration of 2×105 cells/ml in the presence or absence of 10 μM oridonin for 24 h. A total of 200,000 cells were stained with annexin-V and Propidium Iodide (PI) according to the manufacturer’s instructions (BD Biosciences, San Diego, CA). The percentage of annexin-V and Propidium Iodide (PI)-positive cells was analyzed using the BD FACS Calibur platform (Becton Dickinson, LA).
cDNA synthesis and Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
cDNA synthesis and real-time RT-PCR were conducted as previously described [15]. RPMI8226 and KMS-28PE cells were cultured in the presence or absence of oridonin for 2 h. Total RNA was isolated using an RNeasy Mini Kit (Qiagen Sciences, MD). One microgram of total RNA from each sample was reverse transcribed using first-strand cDNA synthesis kit (Qiagen) and subsequently analyzed using RT-PCR (LightCycler, Roche Applied Science, Indianapolis, IN) according to the manufacturer’s instructions. The primers used to amplify human c-myc were as follows: forward, 5’-CTTCTCTCCGTCCTCGGATTCT-3’; and reverse, 5’-GAAGGTGATCCAGACTCTGACCTT-3’. The primers used to amplify the internal control GAPDH were purchased from Nihon Gene Research Laboratories Inc. (Sendai, Japan). SYBR green (Roche Diagnostics, Indianapolis, IN) was used as the PCR probe, and the ratio of c-myc to GAPDH levels in each sample was calculated.
Immunoblotting
Immunoblotting assays were conducted as previously described [16]. Oridonin-treated RPMI8226 and KMS-28PE cells were washed twice with cold phosphate-buffered saline (PBS), and protein lysates were prepared by incubating cell pellets in ice-cold lysis buffer (0.5% NP-40, 10 mM Tris base, 200 mM NaCl, 10% glycerol, 5 mM NaF, and 0.5 mM sodium orthovanadate, pH 7.4) supplemented with 1% protease inhibitor cocktail (EMD Biosciences, San Diego, CA) for 30 min at 4°C. The supernatants were cleared by centrifugation at 4066×g for 10 min, and the samples were eluted with SDS sample buffer (Pierce, Rockford, IL) for 5 min at 95°C. The samples were separated on 12% Tris-glycine gels (Bio-Rad Laboratories, Hercules, CA) and subsequently transferred to polyvinylidene difluoride membranes (Merck Millipore, Tokyo, Japan). Nuclear and cytoplasmic fractions from each extract were prepared using the Cytoplasmic & Nuclear Protein Extraction Kit (101Bio, Palo Alto, CA) according to the manufacturer’s instructions. Immunoblotting assays were conducted according to the instructions provided by the manufacturers of the antibodies, and the proteins were visualized using HRP-conjugated secondary antibodies and the ECL Prime Western Blotting detection reagent (GE Healthcare, Tokyo, Japan). In some experiments, membranes were incubated in a stripping buffer (Pierce, Rockford, IL) according to the manufacturer’s instructions, before being reprobed with additional primary antibodies.
Proteasome inhibition assay
The proteasome inhibition assay was conducted as previously described [17]. RPMI8226 and KMS-28PE cells were incubated in the absence or presence of oridonin (10 μM), MG-132 (10 μM), or both agents for 2 h. The cells were subsequently analyzed using immunoblotting assays with anti-c-Myc and anti-β-actin antibodies as described above.
Immunofluorescence staining
RPMI8226 cells were seeded on glass dishes (Iwaki, Japan), fixed in 4% paraformaldehyde, and washed with PBS 3 times. The cells were permeabilized with 0.1% Triton-X, blocked in blocking buffer (PBS supplemented with 3% bovine serum albumin), and stained with rabbit polyclonal anti-Fbxw7α. The cells were subsequently incubated with AlexaFluor 488-conjugated anti-rabbit IgG (Thermo Fisher Scientific, Japan). The samples were counterstained with Hoechst 33342 (Dojindo, Japan) and AlexaFluor 555 Phalloidin (ThermoFisher Scientific, Japan). Images were obtained using a fluorescence microscope (BZ-X710, Keyence, Japan).
Statistical analysis
The data are expressed as the mean ± Standard Deviation (SD). Statistical differences were assessed using the Student’s t-test. P-values<0.05 were considered statistically significant.
Results
Oridonin suppresses cell proliferation and survival in MM cell lines
We first examined the effect of oridonin on the proliferation of MM cells. As shown in Figure 1A, oridonin treatment inhibited the proliferation of the four MM cell lines evaluated in a dose-dependent manner (IC50 = 8.24 ± 0.18 μM). To examine the effects of oridonin on proliferation and viability of normal blood cells, PBMCs were cultured in the presence or absence of 10 μM oridonin for 24 h. However, the inhibitory effects on normal PBMCs were not observed (data not shown). Next, we examined the effect of oridonin on cell death in MM cells by measuring the frequency of cell death using flow cytometry analysis of cells cultured in the presence or absence of 10 μM oridonin for 24 h and stained with annexin-V and PI. As shown in Figure 1B, the percentage of annexin V+/PI- and annexin- V+/ PI+ cells was greater in oridonin-treated cells compared with control cells. These results suggested that oridonin suppressed proliferation and induced cell death in MM cell lines.
Oridonin suppresses c-Myc expression and activates caspase-3 and PARP in RPMI8226 and KMS-28PE cells
To elucidate the mechanisms by which oridonin suppressed proliferation and induced cell death in MM cells, we examined levels of c-Myc and the anti-apoptotic protein Mcl-1 in RPMI8226 and KMS-28PE cells. As shown in Figure 2A, c-Myc and Mcl-1 protein levels were suppressed in RPMI8226 cells treated with oridonin for 12 h. In contrast, oridonin suppressed c-Myc levels but not Mcl-1 levels in KMS-28PE cells. Oridonin also activated caspase-3 and PARP in both RPMI8226 and KMS-28PE cells. In addition, procaspase-9 levels decreased after 12 h of treatment with oridonin in RPMI8226 cells but not in KMS-28PE cells. These results suggested that oridonin induced caspase-dependent apoptosis in RPMI8226 and KMS-28PE MM cells, however the precise mechanism of apoptosis in each cell line might have differed.
Oridonin does not suppress IRF-4 and Fbxw7α in MM cells
We also examined the effects of oridonin on IRF-4 protein levels, as IRF-4 plays a crucial role in the pathogenesis of MM by directly transactivating the c-myc gene, thereby initiating an autoregulatory feedback loop in MM [18]. As shown in Figure 2B, c-Myc protein levels were suppressed after 2 h of treatment with oridonin. In contrast, IRF-4 protein levels were unaffected, even after 6 h of treatment. As a recent report demonstrated that oridonin enhances Fbxw7α levels and induces apoptosis in leukemia cells by promoting the proteasomal degradation of c-Myc[10], we examined the effects of oridonin on Fbxw7α levels in MM cells. As shown inFigures 2A and2B and, oridonin did not enhance Fbxw7α protein levels in MM cells.
Oridonin induces the downregulation of c-myc mRNA and proteasomal degradation of c-Myc in RPMI8226 and KMS-28PE cells
To elucidate the mechanism of oridonin induced suppression of c-Myc protein levels, we conducted a proteasome inhibition assay. As shown in Figure 3A treatment with the proteasome inhibitor MG132 blocked oridonin-mediated downregulation c-Myc expression in RPMI8226 and KMS-28PE cells, suggesting that c-Myc might be degraded via the ubiquitin-proteasome pathway. Next, we examined the effect of oridonin on the transcription of the c-myc oncogene. As shown in Figure 3B, quantitative RT-PCR analysis demonstrated that c-myc mRNA levels decreased after 2 h of treatment with 10 μM oridonin in RPMI8226 and KMS-28PE cells. These results indicated that oridonin might suppress c-Myc protein levels via both transcriptional repression and proteasomal degradation.
Oridonin induces nuclear translocation of Fbxw7α in RPMI8226 cells
Next, we investigated the mechanism by which oridonin promotes c-Myc degradation in MM cells. c-Myc is a target of Fbxw7α, a component of the SCF-like ubiquitin ligase complex [19]. As oridonin did not alter Fbxw7α expression at the protein level in MM cells (Figures 2A and2B and), we hypothesized that oridonin might affect the intracellular localization of Fbxw7α. Therefore, we treated RPMI8226 cells with oridonin and evaluated the nuclear localization of Fbxw7α using immunoblotting (Figure 4A). Nuclear levels of Fbxw7α increased, whereas nuclear levels of c-Myc decreased after 4 h of oridonin treatment. In contrast, nuclear levels of the HDAC1 loading control were unaffected by oridonin treatment. Moreover, fluorescence immunostaining assays demonstrated that oridonin induced Fbxw7α accumulation in the nucleus Figure 4B). These results suggested that oridonin might trigger the nuclear translocation of Fbxw7α, thereby promoting the proteasomal degradation of c-Myc.
Discussion
In this study, we demonstrated that oridonin suppressed the proliferation of MM cells in a dose-dependent manner and induced apoptosis. In addition, oridonin activated caspase-3 and PARP, indicating that oridonin induced caspase-dependent apoptosis in MM cells. However, oridonin induced a relative slow activation of these proteins in KMS-28PE compared to RPMI8226. KMS-28PE cells have a high-risk cytogenetic feature such as t(4;14) [13]. In contrast, RPMI8226 cells have the standard-risk feature such as hyperdiploidy [12]. On the other hand,oridonin suppressed Mcl-1 and procaspase-9 levels in RPMI8226 but not in KMS-28PE cells. Similarly, Ikezoe et al. demonstrated that oridonin down regulated Mcl-1 and Bcl-xL levels in RPMI8226 and adult T-cell leukemia (MT-1) cells [9]. A recent study demonstrated that oridonin induced apoptosis, increased levels of activated caspase-3 and caspase-9, and decreased mitochondrial membrane potential in gastric cancer cells [20]. In contrast, Kang et al. reported that oridonin activated caspase-8 in human laryngeal cancer cells [21]. These findings suggest that the cell death pathways affected by oridonin might differ according to cell type and cytogenetic features.
We also found that oridonin suppressed c-Myc protein levels in MM cells. Consistent with these findings, a previous study demonstrated that oridonin suppressed c-Myc levels in K562 cells [10]. c-Myc overexpression is an oncogenic event characteristic of MM [5,22]. A previous study reported that c-Myc was overexpressed in 40 % of MM patients [23]. In addition, c-Myc activation is detectable in 67 % of MM patients, and is associated with hyper diploidy and shorter survival time [5]. Walker et al. recently reported that MM patients with c-myc translocations had lower progressionfree and overall survival rates [24]. These findings suggest that c-Myc overexpression and/or activation is associated with a poor prognosis in MM. Holien et al. demonstrated that MM cells depend on c-Myc for survival, as the ablation of c-myc using small hairpin RNAs or small molecule inhibitors induced MM cell death [6]. In addition, recent reports have shown that the BET inhibitor JQ1 suppressed cell proliferation and c-Myc levels [7,25]. Together, these findings suggest that c-Myc represents a potential target of MM treatment. Unfortunately, despite our ever growing understanding of the biology of c-myc, targeted clinical therapeutic strategies for tumors with enhanced c-myc expression are not yet available. The findings of the present study clearly illustrate that oridonin suppressed c-Myc in MM cells. A recent study identified a positive auto regulatory loop between IRF-4 and c-Myc during normal B-cell activation as well as in MM [18]. We found that oridonin did not suppress IRF-4 levels in RPMI8226 or KMS-28PE cells, indicating that the effects of oridonin are specific to c-Myc.
c-Myc is tightly regulated at multiple biologic levels, and it is regulated by Fbxw7 at the posttranslational level [19,26]. Fbxw7, a component of the ubiquitin-proteasome system, is an E3 ubiquitin ligase that targets c-Myc. Huang et al. demonstrated that oridonin suppressed c-Myc protein levels and that this effect was mediated by Fbxw7 [10]. In addition, Fbxw7 was rapidly upregulated in K562 cells and other leukemia and lymphoma cells treated with oridonin. These findings prompted us to examine if oridonin upregulates Fbxw7α in MM cells. However, we did not observe an increase in Fbxw7α levels in oridonin-treated MM cells. E3 ubiquitin ligases catalyze protein degradation via the ubiquitin-proteasome system, and their activity is tightly controlled. One level of regulation involves their subcellular localization. The Fbxw7 gene encodes three mRNAs with unique 5’ exons and 10 common exons [27]. The three Fbxw7 protein isoforms share known functional domains and differ in their localization. Fbxw7α is nuclear, Fbxw7β is cytoplasmic, and Fbxw7γ is predominantly nucleolar [28]. Fbxw7α is the most abundant isoform and accounts for the bulk of known Fbxw7 functions [29]. Therefore, we examined if oridonin affects the subcellular localization of Fbxw7α in MM cells. Interestingly, we found that oridonin induced the nuclear accumulation of Fbxw7α in RPMI8226 cells. Furthermore, oridonin-mediated suppression of c-Myc protein levels was reversed by the proteasome inhibitor MG132. These findings suggest that oridonin might suppress c-Myc levels, at least in part, by promoting c-Myc protein degradation via the nuclear accumulation of Fbxw7α in MM cells. King et al. showed that Fbxw7α mutations specifically affect the ubiquitylation and half-life of c-Myc protein, a key T cell acute lymphoblastic leukemia oncogene [30]. The authors also demonstrated that R465C mutation renders Fbxw7 incapable of ubiquitylating c-Myc. We did not examine the ubiquitylation of c-Myc in oridonin-treated MM cells in this study. On the other hand, Durgan et al. demonstrated that phosphorylation of Fbxw7α at Ser10 can regulate its nuclear localization in mutational analyses [31]. Because cancer-associated mutations in nearby residues influence Fbxw7α localization, there is the possibility of presence of such mutations within Fbxw7α gene in MM cells. To clarify the precise mechanism by which oridonin induces the degradation of c-Myc in MM cells, further studies are needed. We observed that c-myc mRNA levels slightly decreased in MM cells treated with oridonin for 2 h. These observations indicated that oridonin reduced c-Myc expression mainly through the proteasomal degradation rather than the transcriptional repression. However, the precise mechanism mediating the effects of oridonin on c-Myc remains unclear and warrants further investigation in future studies.
In conclusion, oridonin induces caspase-dependent apoptosis in MM cells. In addition, oridonin suppresses the critical MMassociated oncoprotein c-Myc by promoting c-myc transcriptional repression and inducing c-Myc protein degradation via the nuclear accumulation of Fbxw7α. Our data indicate that clinical studies of oridonin in patients with MM may be warranted.
References
- Kumar SK, Rajkumar SV, Dispenzieri A, Lacy MQ, Hayman SR, et al. (2008) Improved survival in multiple myeloma and the impact of novel therapies. Blood 111: 2516-2520.
- Kumar SK, Dispezieri A, Lacy MQ, Gertz MA, Baudi FK, et al. (2014) Continued improvement in survival in multiple myeloma: Changes in early mortality and outcomes in older patients. Leukemia 28: 1122-1128.
- Dang CV (2012) MYC on the path to cancer. Cell 149: 22-35.
- Delgado MD, Leon J (2010) Myc roles in hematopoiesis and leukemia. Genes Cancer 1: 605-616.
- Chng WJ, Huang GF, Chung TH, Ng SB, Gonzalez-Paz N, et al. (2011) Clinical and biological implications of MYC activation: A common difference between MGUS and newly diagnosed multiple myeloma. Leukemia 25: 1026- 1035.
- Holien T, Vatsveen TK, Hella H, Waage A, Sundan A, et al. (2012) Addiction to c-MYC in multiple myeloma. Blood 120: 2450-2453.
- Asahi M, Ito S, Takano M, Ishida Y (2016) YM155 suppresses proliferation and survival 0f multiple myeloma cells via proteasomal degradation of c-Myc. J Med Oncol Ther 1: 62-71.
- Ikezoe T, Chen SS, Heber D, Taguchi H, Koeffler HP (2001) Baicalin is a major component of PC-SPES which inhibits the proliferation of human cancer cells via apoptosis and cell cycle arrest. Prostate 49: 285-292.
- Ikezoe T, Yang Y, Bandobashi K, Saito T, Takemoto S, et al. (2005) Oridonin, a diterpenoid purified from Rabdosia rubescens, inhibits the proliferation of cells from lymphoid malignancies in association with blockade of the NFkappa B signal pathways. Mol Cancer Ther 4: 578-586.
- Huang HL, Weng HY, Wang LQ, Yu CH, Huang QJ, et al. (2012) Triggering Fbw7-mediated proteasomeal degradation of c-Myc by oridonin induces cell growth inhibition and apoptosis. Mol Cancer Ther 11: 1155-1165.
- Zeng R, Chen Y, Zhao S, Cui GH (2012) Autophagy counteracts apoptosis in human multiple myeloma cells exposed to oridonin in vitro via regulating intracellular ROS and SIRT1. Acta Phamacol Sin 33: 91-100.
- Matsuoka Y, Moore GE, Yagi Y, Pressman D (1967) Production of free light chains of immunoglobulin by a hematopoietic cell line derived from a patient with multiple myeloma. Proc Soc Exp Med 125: 1246-1250.
- Otsuki T, Yata K, Sakaguchi H, Kurebayashi J, Matsuo Y, et al. (2002) Interleukin 10 abolishes the growth inhibitory effects of all-trans retinoic acid on human myeloma cells. Br J Haematol 116: 787-795.
- Miura Y, Tsujioka T, Nishimura Y, Sakaguchi H, Maeda M, et al. (2006) TRAIL expression up-regulated by interferon-gamma via phosphorylation of STAT1 induces myeloma cell death. Anticancer Res 26: 4115-4124.
- Sasaki R, Ito S, Asahi M, Ishida Y (2015) YM155 suppresses cell proliferation and induces cell death in human adult T-cell leukemia/lymphoma cells. Leuk Res 39: 1473-1479.
- Ito S, Mantel CR, Han MK, Basu S, Fukuda S, et al. (2007) Mad2 is required for optimal hematopoiesis: Mad2 associates with c-Kit in MO7e cells. Blood 109: 1923-1930.
- Ito S, Oyake T, Murai K, Ishida Y (2010) Deguelin suppresses cell proliferation via the inhibition of survivin expression and STAT3 phosphorylation in HTLV- 1-transformed T cells. Leuk Res 34: 352-357.
- Shaffer AL, Emre NC, Lamy L, Ngo VN, Wright G, et al. (2008) IRF4 addiction in multiple myeloma. Nature 454: 226-231.
- Yada M, Hatakeyama S, Kamura T, Nishiyama M, Tsunematsu R, et al. (2004) Phosphorylation-dependent degradation of c-Myc is mediated by the F-box protein Fbw7. EMBO J 23: 2116-2125.
- Shi M, Lu XJ, Zhang J, Diao H, Li G, et al. (2016) Oridonin, a novel lysine acetyltransferases inhibitor, inhibits proliferation and induces apoptosis in gastric cancer cells through p53- and caspase-3-mediated mechanisms. Oncotarget 7: 22623-22631.
- Kang N, Cao SJ, Zhou Y, He H, Tashiro S, et al. (2015) Inhibition of caspase-9 by oridonin, a diterpenoid isolated from Rabdosia rubescens, augments apoptosis in human laryngeal cancer cells. Int J Oncol 47: 2045-2056.
- Kuehl WM, Bergsagel PL (2012) MYC addiction: a potential therapeutic target in MM. Blood 120: 2351-2352.
- Szabo AG, Gang AO, Pedersen MØ, Poulsen TS, Klausen TW, et al. (2016) Overexpression of c-myc is associated with adverse clinical features and worse overall survival in multiple myeloma. Leuk Lymphoma 57: 2526-2534.
- Walker BA, Wardell CP, Brioli A, Boyle E, Kaiser MF, et al. (2014) Translocations at 8q24 juxtapose MYC with genes that harbor supermenhancers resulting in overexpression and poor prognosis in myeloma patients. Blood Cancer J 4: e191.
- Delmore JE, Issa GC, Lemieux ME, Rahl PB, Shi J, et al. (2011) BET bromodomain inhibition as a therapeutic strategy to target c-Myc. Cell 146: 904-917.
- Welcker M, Orian A, Jin J, Grim JE, Harper JW, et al. (2004) The Fbw7 tumor suppressor regulates glycogen synthase kinase 3 phosphorylationdependent c-Myc protein degradation. Proc Natl Acad Sci U S A 101: 9085- 9090.
- Spruck CH, Strohmaier H, Sangfelt O, Muller HM, Hubalek M, et al. (2002) hCDC4 gene mutations in endometrial cancer. Cancer Res 62: 4535-4539.
- Welcker M, Orian A, Grim JE, Eisenman RN, Clurman BE (2004) A nucleolar isoform of the Fbw7 ubiquitin ligase regulates c-Myc and cell size. Curr Biol 14: 1852-1857.
- Grim JE, Gustafson MP, Hirata RK, Hagar AC, Swanger J, et al. (2008) Isoform- and cell cycle-dependent substrate degradation by the Fbw7 ubiquitin ligase. J Cell Biol 181: 913-920.
- King B, Trimarchi T, Reavie L, Xu L, Mullenders J, et al. (2013) The ubiquitin ligase FBXW7 modulates leukekmia-initiating cell activity by regulating MYC stability. Cell 153: 1552-1566.
- Durgan J, Parker PJ (2010) Regulation of the tumor suppressor Fbw7α by PKC-dependent phosphorylation and cancer-associated mutations. Biochem J 432: 77-87.