
Review Article
*Address for Correspondence: Larry S. Schlesinger, The Ohio State University, 798 Biomedical Research Tower, 460 W. 12th Avenue, Columbus, OH 43210, USA, Tel: (614) 292-8789; Fax: (614)-292-9616; E-mail: larry.schlesinger@osumc.eduAbul K. Azad, The Ohio State University, 711 Biomedical Research Tower, 460 W. 12th Avenue, Columbus, OH 43210, USA, Tel: (614) 292-4635; Fax: (614)-292-9616; E-mail: abul.azad@osumc.edu
Citation: Azad AK, Rajaram MVS, Schlesinger LS. Exploitation of the Macrophage Mannose Receptor (CD206) in Infectious Disease Diagnostics and Therapeutics. J Cytol Molecul Biol. 2014;1(1): 5.
Copyright © 2014 Azad AK, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use,distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Cytology & Molecular Biology | ISSN: 2325-4653 | Volume: 1, Issue: 1
Submission: 28 November 2013 | Accepted: 06 January 2014 | Published: 10 January 2014
Reviewed & Approved by: Dr. Sankar N. Maity, Department of Genitourinary Medical Oncology, University of Texas MD Anderson Cancer Center, USA

Exploitation of the Macrophage Mannose Receptor (CD206) in Infectious Disease Diagnostics and Therapeutics
Abul K. Azad*,Murugesan V. S. Rajaram and Larry S. Schlesinger*
- Department of Pathology, University of Patras School of Medicine, 265 04 Rion, Patras, Greece
- Center for Microbial Interface Biology and the Department of Microbial Infection and Immunity, The Ohio State University, USA
*Address for Correspondence: Larry S. Schlesinger, The Ohio State University, 798 Biomedical Research Tower, 460 W. 12th Avenue, Columbus, OH 43210, USA, Tel: (614) 292-8789; Fax: (614)-292-9616; E-mail: larry.schlesinger@osumc.eduAbul K. Azad, The Ohio State University, 711 Biomedical Research Tower, 460 W. 12th Avenue, Columbus, OH 43210, USA, Tel: (614) 292-4635; Fax: (614)-292-9616; E-mail: abul.azad@osumc.edu
Citation: Azad AK, Rajaram MVS, Schlesinger LS. Exploitation of the Macrophage Mannose Receptor (CD206) in Infectious Disease Diagnostics and Therapeutics. J Cytol Molecul Biol. 2014;1(1): 5.
Copyright © 2014 Azad AK, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use,distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Cytology & Molecular Biology | ISSN: 2325-4653 | Volume: 1, Issue: 1
Submission: 28 November 2013 | Accepted: 06 January 2014 | Published: 10 January 2014
Reviewed & Approved by: Dr. Sankar N. Maity, Department of Genitourinary Medical Oncology, University of Texas MD Anderson Cancer Center, USA
Abstract
The macrophage mannose receptor (MR, CD206) is a C-type lectin expressed predominantly by most tissue macrophages, dendritic cells and specific lymphatic or endothelial cells. It functions in endocytosis and phagocytosis, and plays an important role in immune homeostasis by scavenging unwanted mannoglycoproteins. More attention is being paid to its particularly high expression in tissue pathology sites during disease such the tumor microenvironment. The MR recognizes a variety of microorganisms by their mannan-coated cell wall, which is exploited by adapted intracellular pathogens such as Mycobacterium tuberculosis, for their own survival. Despite the continued development of drug delivery technologies, the targeting of agents to immune cells, especially macrophages, for effective diagnosis and treatment of chronic infectious diseases has not been addressed adequately. In this regard, strategies that optimize MR-mediated uptake by macrophages in target tissues during infection are becoming an attractive approach. We review important progress in this area.Introduction
Infectious diseases account for more than 25% of human deaths in the world annually and are currently the third leading cause of death in the United States [1,2]. The optimism in the early 1970s of eradicating infectious diseases with the discovery of powerful antibiotics, vaccines, and antiviral drugs has been lost with the emergence of multiple anti-infective resistant microbes. As infectious diseases remain very significant health problems at home and around the world, a central component of the world-wide effort to eliminate or thwart these diseases will rest on having markedly improved imaging/diagnostic capabilities as well as more effective therapies and vaccines. The lack of accessibility of many diagnostic and chemotherapeutic agents to infected or diseased tissue sites in patients with a variety of infectious diseases remains a formidable obstacle to effective cures, especially for chronic and recurring infections due to intracellular pathogens of macrophages. Despite the continued development of drug delivery technologies, the targeting of agents to immune cells, especially macrophages, for effective diagnosis and treatment of the underlying diseases has not been addressed adequately. Through ongoing efforts in the Center for Microbial Interface Biology at The Ohio State University, attention is being paid to the mannosylation of clinical agents for advanced imaging, diagnostics and therapeutic strategies that optimize MR-mediated uptake by macrophages in target tissues during infection. It is anticipated that such strategies will also enable the use of smaller amounts of agents for optimal clinical effects, thereby increasing efficacy and reducing toxicity.Macrophage Mannose Receptor
The mannose receptor (MR, CD206 or MRC1) is a transmembrane glycoprotein that belongs to the C-type lectin family and is expressed predominantly by most tissue macrophages, dendritic cells (DCs) and selected lymphatic or liver endothelial cells [3]. Its extracellular regions include an N-terminal cysteine-rich (CR) domain that binds glycoproteins bearing sulfated sugars that terminate in 4-SO4GalNAc, a fibronectin II (FNII) domain, and eight carbohydrate recognition domains (CRDs) that bind sugars like mannose and fucose with high affinity. The MR serves as a homeostatic receptor by binding and scavenging unwanted high mannose N-linked glycoproteins as well as pituitary hormones from the circulation [4-6]. Since many pathogenic microbes are coated with mannose-containing structures, the macrophage MR interacts with those pathogens in a form of the host molecular mimicry [7].MR and Pathogen Recognition
The MR recognizes a variety of microorganisms, including bacteria, fungi, virus, and parasites. Notably, Mycobacterium tuberculosis, Streptococcus pneumonia, Yersinia pestis, Candida albicans, Pneumocystis carinii, Cryptococcus neoformans, HIV, influenza virus, dengue virus, and Leishmania species bind to the MR and are endocytosed by macrophages [5,8]. Many MR-binding organisms are intracellular pathogens. Since the MR is an abundantly expressed endocytic, recycling receptor [9], targeting this receptor is a viable and attractive strategy for the delivery of carbohydrate containing imaging/diagnostic agents as well as the intracellular delivery of therapeutics for many infectious diseases.Rationale for MR Targeting
The lack of accessibility of many diagnostic and chemotherapeutic agents in infected or diseased sites of patients with diseases like cancer and infectious diseases has remained a clinical challenge. Despite the continued development of drug delivery technologies, the effective targeting of drugs to macrophages for the diagnosis and treatment of the underlying diseases remains to be proven. Based on a growing literature, the feasibility that mannosylation of imaging agents, diagnostics and/or therapeutics will lead to clinically relevant MR-mediated uptake by macrophages in target tissues or organs is much increased. Furthermore, enhanced uptake is predicted to require smaller doses of the agents sufficient for optimal clinical effects, thereby reducing the toxicity of administered substances.Strategies for Small Molecule Delivery to Macrophages
For effective and targeted delivery, small molecule (e.g., drug) carriers such as liposomes, microparticles, nanoparticles, dendrimers, etc. are used. Most of the carrier molecules are inert and exhibit multiple advantages of biocompatibility, biodegradability, high charge density and non-toxicity. In most cases, these carriers are functionalized with mannose (mannosylated) or with anti-MR nanobody to use as substrates for targeting the MR. The small molecule carriers are able to traverse the different membrane barriers of macrophages and release their cargo inside the cells. The same principle has been exploited recently by using mannosylated nanoparticles (polymeric micelles) for high efficiency delivery of MR-targeted siRNA into macrophages [10]. The MR has recently been shown to mediate the uptake and endosomal delivery of the intracellular TLR9 ligand CpG motif-containing oligodeoxynucleotides in murine peritoneal macrophages without using any mannosylated carriers [11]. More advanced delivery systems targeting the macrophage MR have also been reported in recent years in various disease models [12-14]. Singodia et al. used an alternate approach to target the macrophage MR by coating antibiotic-loaded liposomes with a sulfated sugar ligand of the receptor (4-SO4GalNAc) rather than mannoses, which led to enhanced antibiotic uptake by the CR domain of the MR and thus opens up a new drug delivery strategy via this receptor [12]. Two novel polyethylene glycol-treated (PEGylated) nanocarriers have been developed for high efficiency MR-mediated macrophage targeting: one by using PEG-sheddable, mannose-functionalized nanoparticles in the acidic tumor microenvironment [13] and the other by incorporating PEG and peptide in a dendritic structure device known as “PEGtide dendrons” functionalized with mannose residues [14]. Because of their modular design, PEGtide dendrons were suggested to be advanced carriers in imaging and drug delivery applications.Targeting the MR to Tumor-Associated Macrophages (TAMs) in Cancer
Tissue macrophages, the major patrolling immune cells of the innate immune system as well as the communication cells for the development and maintenance of the adaptive immune system, represent a highly diverse population in terms of phenotype and function depending on their tissue sites and state of activation [15]. They have variably been termed M1, M2, immunoregulatory, etc. highlighting their spectrum of activities. Alveolar macrophages are prototypic alternatively activated macrophages and express abundant MR which mediates interaction with various air-borne pathogens [5]. In the cancer tissue microenvironment, tumor associated macrophages or TAMs have been described which play important roles in tumor invasion, proliferation and metastasis [16]. These are M2-type macrophages and express high levels of the MR [17]. Studies in cancer research are targeting the MR expressed on TAMs. These studies are providing proof of principle that the MR is an appealing bio-target for cancer diagnosis and therapy. Mannose-coated liposomes have been used for imaging of TAMs by positron emission tomography (PET) [18], and to mediate MR-directed anti-cancer drug delivery in a mouse model of peritoneal metastasis and in human gastric cancer [19]. Using single-photon emission computed tomography/micro-computed tomography (SPECT/micro-CT) has enabled in vivo imaging of TAMs by nanobody-based targeting of the MR in mouse models of lung and breast cancer [20]. A similar approach has been recently applied to autoimmune disease where a SPECT/micro-CT imaging of joint inflammation was performed by targeting nanobody-based macrophage MR in a mouse model of rheumatoid arthritis [21]. Thus, TAMs represent an appealing bio-target for cancer imaging, diagnosis and therapy. The successful optimization of techniques for efficient MR-mediated delivery of imaging and therapeutic agents in these types of studies set the stage for targeting MR-expressing macrophages for similar types of clinical applications in infectious diseases including tuberculosis (TB).Targeting the MR in Infectious Diseases
TB as a modelAs noted above, many pathogens are surface-coated with specific carbohydrates that are recognized by the macrophage MR and other C-type lectins. Research is beginning to exploit the strategy of receptor and ligand interaction, and make use of mannosylated “pathogen-like” polyanhydride nanoparticles for delivery of antigens into macrophages and DCs, the latter being another critical immune cell in innate and adaptive immunity [22,23]. Liposomes are often used as a carrier system for antimicrobial agents and, in general, to improve their bio-distribution, reduce cell toxicity, and increase the therapeutic index at the infection site [24]. In this regard, the MR represents a potential target for efficient delivery of mannosyated liposomes.TB is a respiratory infection caused by the bacterial pathogen Mycobacterium tuberculosis. It currently infects approximately one third of the world’s population and is the second leading cause of death globally due to a single infectious agent with ongoing worldwide efforts to improve therapies and vaccines [25]. M. tuberculosis is a prototypic intracellular pathogen of macrophages which play a major role in both latent and active TB. Macrophages are an essential component for granuloma formation and maintenance. The granuloma is where M. tuberculosis is controlled and persists, yet this unique environment remains one of the least understood aspects of the host-pathogen relationship [26]. What is widely recognized, however, is that the granuloma microenvironment represents a formidable barrier to the delivery of diagnostic agents and therapies, akin to the tumor microenvironment, and some parallels can be drawn, including physiological barriers such as reduced oxygen tension and altered phenotype and function of macrophages [27]. We currently lack the ability to accurately image granulomas in patients with latency, a condition in which treatment can reduce the risk of developing active TB.
Targeting the macrophage MR is a potential and an attractive strategy for the imaging, diagnosis and therapy of TB (Figure 1). Our group discovered the role of the MR in the phagocytosis of M. tuberculosis by human macrophages 20 years ago [28], and, more recently, MR’s role in regulating macrophage responses to this pathogen [29,30]. To date, there has been no report on the systematic evaluation of the MR on macrophages within TB granulomas akin to TAMs. However, it is likely that such macrophages express the MR (CD206) in addition to CD163 [27]. The MR has been implicated in macrophage adhesion and fusion during granuloma formation [31]. In addition, PPARγ mediates induction of the MR and foamy cells, the latter found in granulomas [32]. We have found that PPARγ is up-regulated by M. tuberculosis engagement of the MR [30] which could potentially help sustain the regulated inflammatory environment within granulomas [27]. Since the TB granuloma provides a tangible barrier to antibiotic penetration [33] and the MR is predicted to be abundantly expressed on macrophages, foamy cells and DCs, all cells being present in the granulomas, the MR could be an attractive target for imaging agents and drug delivery systems in this microenvironment akin to strategies being developed in the cancer field (Figure 2). In a recent study, D’Addio et al. optimized macrophage MR-specific targeting for anti-TB drugs by using nanocarriers functionalized with polymannoses and found that maximum macrophage uptake of the drugs occurred with increased mannose density on nanocarrier surfaces [34].
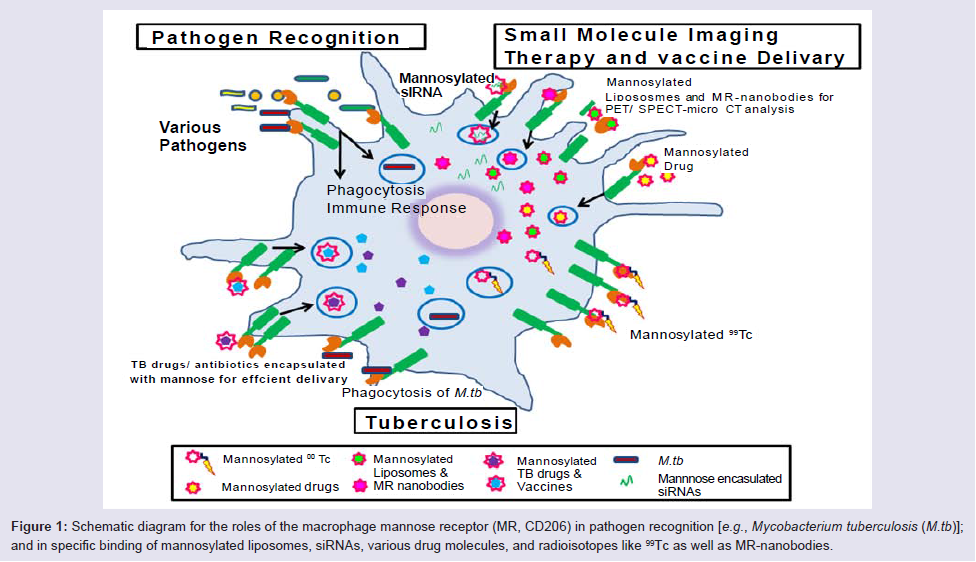
Figure 1: Schematic diagram for the roles of the macrophage mannose receptor (MR, CD206) in pathogen recognition [e.g., Mycobacterium tuberculosis (M.tb)]; and in specific binding of mannosylated liposomes, siRNAs, various drug molecules, and radioisotopes like 99Tc as well as MR-nanobodies.
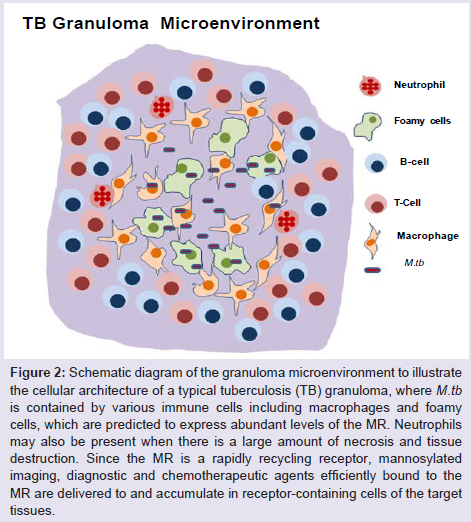
Figure 2: Schematic diagram of the granuloma microenvironment to illustrate the cellular architecture of a typical tuberculosis (TB) granuloma, where M.tb is contained by various immune cells including macrophages and foamy cells, which are predicted to express abundant levels of the MR. Neutrophils may also be present when there is a large amount of necrosis and tissue destruction. Since the MR is a rapidly recycling receptor, mannosylated imaging, diagnostic and chemotherapeutic agents efficiently bound to the MR are delivered to and accumulate in receptor-containing cells of the target tissues.
Targeting the MR for Vaccine Delivery
The macrophage MR can mediate the presentation of mycobacterial antigens to T cells in the development of an adaptiveimmune response [35].This property raises the potential for targeting the MR and other C-type lectins in the development of effective vaccines [36]. In this context, several studies provide evidence that the MR pathway can be targeted for vaccine delivery [37-39]. For example, a novel DNA vaccine formulation enhances cytotoxic T-lymphocyte activity through efficient gene delivery to DCs by MR-mediated endocytosis [40]. The MR endocytic pathway can also be used to deliver DNA-based vaccines into antigen-presenting cells using mannosylated liposomes [41]. The use of cationic mannosylated liposomes complexed with plasmid DNA has shown high transfection efficiency due to recognition by the MR both in vitro and in vivo [42]. The concept of T-cell activation by this approach is supported by a study where CD8+ T-cell activation was successfully induced via the MR-mediated endocytic pathway providing further evidence for the use of the MR in vaccine therapy for TB and other infectious diseases [43].Concluding Remarks
The majority of strategies for imaging, diagnostic or therapeutic agents that target the macrophage MR have used mannosylation of their carriers rather than modifying the agents themselves. Incorporation of polymannoses in the structures of the agents could be an effective approach for their faster, more penetrable delivery to target sites, assuming the compounds remain non-toxic and do not lose their efficacy upon mannosylation. For example, the FDA recently approved technetium (99Tc)-labeled tilmanocept from Navidea Pharmaceuticals as the first receptor-targeted poly mannosylated radiopharmaceutical [44] used for precision imaging of sentinel lymph nodes in solid tumor staging. Targeting the MR by tilmanocept and derived compounds has the potential to enhance imaging agents, particularly within TB granulomas, as well as the delivery of chemotherapeutic agents (Figure 1 and Figure 2). Innovative strategies for targeting the MR pathway for the rational design and effective delivery of agents into uniquely located tissue macrophages hold promise for the diagnosis and treatment of a variety of infectious diseases.Acknowledgements
The work discussed from our lab in this review was supported in part by the NIH grants awarded to LSS (R01 AI059639 and R01 AI052458).References
- WHO (2012) Global Tuberculosis Report 2012.
- Fauci AS, Touchette NA, Folkers GK (2005) Emerging infectious diseases: a 10-year perspective from the National Institute of Allergy and Infectious Diseases. Emerg Infect Dis 11: 519-525.
- Taylor PR, Martinez-Pomares L, Stacey M, Lin HH, Brown GD, et al. (2005) Macrophage receptors and immune recognition. Annu Rev Immunol 23: 901-944.
- Martinez-Pomares L, Linehan SA, Taylor PR, Gordon S (2001) Binding properties of the mannose receptor. Immunobiology 204: 527-535.
- Allavena P, Chieppa M, Monti P, Piemonti L (2004) From pattern recognition receptor to regulator of homeostasis: the double-faced macrophage mannose receptor. Crit Rev Immunol 24: 179-192.
- Lee SJ, Evers S, Roeder D, Parlow AF, Risteli J, et al. (2002) Mannose receptor-mediated regulation of serum glycoprotein homeostasis. Science 295: 1898-1901.
- Medzhitov R (2007) Recognition of microorganisms and activation of the immune response. Nature 449: 819-826.
- Taylor PR, Gordon S, Martinez-Pomares L (2005) The mannose receptor: linking homeostasis and immunity through sugar recognition. Trends Immunol 26: 104-110.
- Stahl P, Schlesinger PH, Sigardson E, Rodman JS, Lee YC (1980) Receptor-mediated pinocytosis of mannose glycoconjugates by macrophages: characterization and evidence for receptor recycling. Cell 19: 207-215.
- Yu SS, Lau CM, Barham WJ, Onishko HM, Nelson CE, et al. (2013) Macrophage-Specific RNA Interference Targeting via “Click”, Mannosylated Polymeric Micelles. Mol Pharm 10: 975-987.
- Moseman AP, Moseman EA, Schworer S, Smirnova I, Volkova T, et al. (2013) Mannose Receptor 1 Mediates Cellular Uptake and Endosomal Delivery of CpG-Motif Containing Oligodeoxynucleotides. J Immunol 191: 5615-5624.
- Singodia D, Verma A, Verma RK, Mishra PR (2012) Investigations into an alternate approach to target mannose receptors on macrophages using 4-sulfated N-acetyl galactosamine more efficiently in comparison with mannose-decorated liposomes: an application in drug delivery. Nanomedicine8: 468-477.
- Zhu S, Niu M, O’Mary H, Cui Z (2013) Targeting of Tumor-Associated Macrophages Made Possible by PEG-Sheddable, Mannose-Modified Nanoparticles. Mol Pharm [Epub ahead of print].
- Gao J, Chen P, Singh Y, Zhang X, Szekely Z, et al. (2013) Novel Monodisperse PEGtide Dendrons: Design, Fabrication, and Evaluation of Mannose Receptor-Mediated Macrophage Targeting. Bioconjug Chem [Epub ahead of print].
- Wynn TA, Chawla A, Pollard JW (2013) Macrophage biology in development, homeostasis and disease. Nature 496: 445-455.
- Mantovani A, Bottazzi B, Colotta F, Sozzani S, Ruco L (1992) The origin and function of tumor-associated macrophages. Immunol Today 13: 265-270.
- Allavena P, Chieppa M, Bianchi G, Solinas G, Fabbri M, et al. (2010) Engagement of the mannose receptor by tumoral mucins activates an immune suppressive phenotype in human tumor-associated macrophages. Clin Dev Immunol 2010: 547179.
- Locke LW, Mayo MW, Yoo AD, Williams MB, Berr SS (2012) PET imaging of tumor associated macrophages using mannose coated 64Cu liposomes. Biomaterials 33: 7785-7793.
- Matsui M, Shimizu Y, Kodera Y, Kondo E, Ikehara Y (2010) Targeted delivery of oligomannose-coated liposome to the omental micrometastasis by peritoneal macrophages from patients with gastric cancer. Cancer Sci 101: 1670-1677.
- Movahedi K, Schoonooghe S, Laoui D, Houbracken I, Waelput W, et al. (2012) Nanobody-based targeting of the macrophage mannose receptor for effective in vivo imaging of tumor-associated macrophages. Cancer Res 72: 4165-4177.
- Put S, Schoonooghe S, Devoogdt N, Schurgers E, Avau A, et al. (2013) SPECT imaging of joint inflammation with Nanobodies targeting the macrophage mannose receptor in a mouse model for rheumatoid arthritis. J Nucl Med 54: 807-814.
- Carrillo-Conde B, Song EH, Chavez-Santoscoy A, Phanse Y, Ramer-Tait AE, et al. (2011) Mannose-functionalized “pathogen-like” polyanhydride nanoparticles target C-type lectin receptors on dendritic cells. Mol Pharm 8: 1877-1886.
- Chavez-Santoscoy AV, Roychoudhury R, Pohl NL, Wannemuehler MJ, Narasimhan B, et al. (2012) Tailoring the immune response by targeting C-type lectin receptors on alveolar macrophages using “pathogen-like” amphiphilic polyanhydride nanoparticles. Biomaterials 33: 4762-4772.
- Naik SR, Desai SK, Shah PD, Wala SM (2013) Liposomes as Potential Carrier System for Targeted Delivery of Polyene Antibiotics. Recent Pat Inflamm Allergy Drug Discov 7: 202-214.
- Zumla A, Nahid P, Cole ST (2013) Advances in the development of new tuberculosis drugs and treatment regimens. Nat Rev Drug Discov 12: 388-404.
- Guirado E, Schlesinger LS, Kaplan G (2013) Macrophages in tuberculosis: friend or foe. Semin Immunopathol 35: 563-583.
- Mattila JT, Ojo OO, Kepka-Lenhart D, Marino S, Kim JH, et al. (2013) Microenvironments in tuberculous granulomas are delineated by distinct populations of macrophage subsets and expression of nitric oxide synthase and arginase isoforms. J Immunol 191: 773-784.
- Schlesinger LS (1993) Macrophage phagocytosis of virulent but not attenuated strains of Mycobacterium tuberculosis is mediated by mannose receptors in addition to complement receptors. J Immunol 150: 2920-2930.
- Kang PB, Azad AK, Torrelles JB, Kaufman TM, Beharka A, et al. (2005) The human macrophage mannose receptor directs Mycobacterium tuberculosis lipoarabinomannan-mediated phagosome biogenesis. J Exp Med 202: 987-999.
- Rajaram MV, Brooks MN, Morris JD, Torrelles JB, Azad AK, et al. (2010) Mycobacterium tuberculosis activates human macrophage peroxisome proliferator-activated receptor gamma linking mannose receptor recognition to regulation of immune responses. J Immunol 185: 929-942.
- McNally AK, DeFife KM, Anderson JM (1996) Interleukin-4-induced macrophage fusion is prevented by inhibitors of mannose receptor activity. Am J Pathol 149: 975-985.
- Peyron P, Vaubourgeix J, Poquet Y, Levillain F, Botanch C, et al. (2008) Foamy macrophages from tuberculous patients’ granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence. PLoS Pathog 4: e1000204.
- Kjellssoni MC, Via LE, Goh A, Weiner D, Low KM, et al. (2012) Pharmacokinetic evaluation of the penetration of antituberculosis agents in rabbit pulmonary lesions. Antimicrob Agents Chemother 56: 446-457.
- D’Addio SM, Baldassano S, Shi L, Cheung L, Adamson DH, et al. (2013) Optimization of cell receptor-specific targeting through multivalent surface decoration of polymeric nanocarriers. J Control Release 168: 41-49.
- Prigozy TI, Sieling PA, Clemens D, Stewart PL, Behar SM, et al. (1997) The mannose receptor delivers lipoglycan antigens to endosomes for presentation to T cells by CD1b molecules. Immunity 6: 187-197.
- Torrelles JB, Azad AK, Henning LN, Carlson TK, Schlesinger LS (2008) Role of C-type lectins in mycobacterial infections. Curr Drug Targets 9: 102-112.
- Keler T, Ramakrishna V, Fanger MW (2004) Mannose receptor-targeted vaccines. Expert Opin Biol Ther 4: 1953-1962.
- Ramakrishna V, Treml JF, Vitale L, Connolly JE, O’Neill T, et al. (2004) Mannose receptor targeting of tumor antigen pmel17 to human dendritic cells directs anti-melanoma T cell responses via multiple HLA molecules. J Immunol 172: 2845-2852.
- Lam JS, Mansour MK, Specht CA, Levitz SM (2005) A model vaccine exploiting fungal mannosylation to increase antigen immunogenicity. J Immunol 175: 7496-7503.
- Hattori Y, Kawakami S, Lu Y, Nakamura K, Yamashita F, et al. (2006) Enhanced DNA vaccine potency by mannosylated lipoplex after intraperitoneal administration. J Gene Med 8: 824-834.
- Lu Y, Kawakami S, Yamashita F, Hashidai M (2007) Development of an antigen-presenting cell-targeted DNA vaccine against melanoma by mannosylated liposomes. Biomaterials 28: 3255-3262.
- Kawakami S, Sato A, Nishikawa M, Yamashita F, Hashida M (2000) Mannose receptor-mediated gene transfer into macrophages using novel mannosylated cationic liposomes. Gene Ther 7: 292-299.
- Burgdorf S, Kautz A, Böhnert V, Knolle PA, Kurtsi C (2007) Distinct pathways of antigen uptake and intracellular routing in CD4 and CD8 T cell activation. Science 316: 612-616.
- Vera DR, Wallace AM, Hoh CK, Mattrey RF (2001) A synthetic macromolecule for sentinel node detection: (99m)Tc-DTPA-mannosyl-dextran. J Nucl Med 42: 951-959