
Research Article
*Address for Correspondence: Habibeh Khoshbouei Ph.D., Department of Neuroscience and Psychiatry, McKnight Brain Institute of the University of Florida School of Medicine, FL 32611, USA, Tel: 35202738115; E-mail: habibeh@ufl.edu
Citation: Butler B, Gamble-George J, Prins P, North A, Clarke JT, Khoshbouei H. Chronic Methamphetamine Increases Alpha-Synuclein Protein Levels in the Striatum and Hippocampus but not in the Cortex of Juvenile Mice. J Addiction Prevention. 2014;2(2): 6.
Copyright © 2014 Butler B, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Addiction & Prevention | ISSN: 2330-2178 | Volume: 2, Issue: 2
Chronic Methamphetamine Increases Alpha-Synuclein Protein Levels in the Striatum and Hippocampus but not in the Cortex of Juvenile Mice
B. Butler, J. Gamble-George, P. Prins, A. North, J.T Clarke and H. Khoshbouei*
- Department of Neuroscience, University of Florida School of Medicine, Gainesville, Fl 32611, USA
*Address for Correspondence: Habibeh Khoshbouei Ph.D., Department of Neuroscience and Psychiatry, McKnight Brain Institute of the University of Florida School of Medicine, FL 32611, USA, Tel: 35202738115; E-mail: habibeh@ufl.edu
Citation: Butler B, Gamble-George J, Prins P, North A, Clarke JT, Khoshbouei H. Chronic Methamphetamine Increases Alpha-Synuclein Protein Levels in the Striatum and Hippocampus but not in the Cortex of Juvenile Mice. J Addiction Prevention. 2014;2(2): 6.
Copyright © 2014 Butler B, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Addiction & Prevention | ISSN: 2330-2178 | Volume: 2, Issue: 2
Submission: 01 December 2014 | Accepted: 19 December 2014 | Published: 22 December 2014
Abstract
Methamphetamine is the second most widely used illicit drug worldwide. More than 290 tons of methamphetamine was synthesized in the year 2005 alone, corresponding to approximately ~3 billion 100 mg doses of methamphetamine. Drug addicts abuse high concentrations of methamphetamine for months and even years. Current reports in the literature are consistent with the interpretation that methamphetamine-induced neuronal injury may render methamphetamine users more susceptible to neurodegenerative pathologies. Specifically, chronic exposure to psychostimulants is associated with increases in striatal alpha-synuclein expression, a synaptic protein implicated in the pathogenesis of neurodegenerative diseases. This raises the question whether methamphetamine exposure affects alpha-synuclein levels in the brain. In this short report, we examined alpha-synuclein protein and mRNA levels in the striatum, hippocampus and cortex of adolescent male mice following a neurotoxic regimen of methamphetamine (24mg/kg/daily/14days). We found that methamphetamine exposure resulted in a decrease in the monomeric form of alpha-synuclein (molecular species 19 kDa) in the striatum and hippocampus, but not in the cortex. Despite the elevation of high molecular weight alpha-synuclein species (>19 kDa), there was no change in the alpha-synuclein mRNA levels in the striatum, hippocampus and cortex of mice exposed to methamphetamine. The methamphetamine-induced increase in high molecular weight alpha-synuclein protein levels might be one of the causal mechanisms or one of the compensatory consequences of methamphetamine-mediated neurotoxicity.Keywords:
Methamphetamine; Parkinson’s disease; Aging; Alphasynuclein; Synaptic transmissionIntroduction
In the United States, oral methamphetamine (Desoxyn, OVATION Pharmaceuticals) is an FDA approved treatment for attention deficit hyperactivity disorder in children and for the short-term treatment of obesity. Methamphetamine is also the second most popular illicit drug worldwide. According to the United Nations Office on Drugs and Crime, more than 290 tons of methamphetamine synthesized in the year 2005 alone [1]. This corresponds to approximately ~3 billion 100 mg doses of methamphetamine. Human methamphetamine users routinely abuse large quantities of the drug (55-640mg) [2,3] for months and even years. Post-mortem brain tissue from methamphetamine users show a decrease in striatal dopamine (DA), dopamine transporter (DAT) density and DA binding affinity [4-6]. Similarly, repeated methamphetamine administration can cause neurotoxicity in primates and rodents [7,8]. The brain tissue of human methamphetamine users exhibits long-term and possibly irreversible damage to DA neurons and loss of striatal DAT even after three years of abstinence [5]. Repeated methamphetamine injections in rats promote formation of higher molecular weight (>150 kDa) DAT complexes 24-48 hours post treatment [9]. This effect is considered as one putative underlying mechanisms responsible for decreasing DAT function and was suppressed by manipulations that prevent the persistent methamphetamine-mediated deficits in dopaminergic function [9].Recent studies [10,11] suggest that methamphetamine exposure during early stages of life may increase risk of developing Parkinsonlike symptoms. The risks of developing Parkinson-like symptoms were shown to rise by 75% in methamphetamine abusers [10]. Other studies suggest that chronic exposure to psychostimulants may increase the level of alpha-synuclein, a synaptic protein implicated in the pathogenesis of neurodegenerative diseases. For example, Mash and colleagues [12-14] reported elevated alpha-synuclein levels in the midbrain DA neurons of chronic cocaine abusers. Western blot and immuno-autoradiography analysis of postmortem brain tissue obtained from chronic cocaine abusers and age-matched drug-free control subjects revealed threefold higher alpha-synuclein levels in the dopaminergic neurons of the substantia nigra and ventral tegmental area of cocaine abusers as compared to controls [12-14]. Furthermore, it has been demonstrated that overexpression of alpha-synuclein within neurons of the substantia nigra induced by amphetamine derivatives is related to the occurrence of intracellular inclusions of this protein [15]. Tavassoly and Lee [16] documented that methamphetamine binds to the N-terminus of alpha-synuclein and causes a conformational change in the protein that can lead to stabilization of the oligomeric form of alpha-synuclein implicated in neurotoxicity induction.
Accordingly, in this short report we tested the hypothesis that adolescent male mice exposed to a chronic neurotoxic regimen of methamphetamine will exhibit elevated alpha-synuclein levels in the brain structures thought to be targets of methamphetamine neurotoxicity.
Materials and Methods
AnimalsThirty six C57BL/6J male mice at 4-5 weeks of age were obtained from the Jackson Laboratory (Bar Harbor, ME, USA). These animals were within the adolescent age range for rodents (postnatal days 28-42) [17,18]. All of the animals were housed in the AALAC accredited animal care facility at Meharry Medical College, under constant temperature and a 12-h light/dark cycle (lights on: 7 a.m-7 p.m), with food and water provided ad libitum. All experimental procedures complied with the NIH Guide for the Care and Use of Laboratory Animals, and were conducted with the approval of Meharry Medical College Institutional Animal Care and Use Committee. All efforts were made to keep the stress and discomfort at a minimum and to decrease the number of animals utilized in this study. The animals were gently handled every day for five days prior to experimentation, and throughout the study.
Drug Treatment
Methamphetamine addicts may abuse large quantities of the drug (55-640mg methamphetamine) over a few hours for months and years [3]. In the United States, arrest reports show methamphetamine blood levels can be above 300 mg/l [2]. To examine methamphetamine induced neurotoxicity in animal models, various non-contingent high doses of methamphetamine have been used. For example, methamphetamine regimens of 60mg/kg/day [19], 40 mg/kg/day [20], 32 mg/kg/day [21], 30mg/kg/day [22-24], and 24 mg/kg/day/14 days [25] have been used to study the neurotoxic consequence of methamphetamine exposure. We selected the daily dose of 24mg/kg/ day methamphetamine for 14 days as described previously [19,22-25] and reported to produce signs of toxicity (North et al. 2013). The animals were gently handled every day for five days prior to experimentation, and received 14 days of intraperitoneal (i.p.) 24mg/ kg/day methamphetamine or saline (N=18 methamphetamine, N=18 saline, half the brains were used for western blot analysis, the other half used for RNA isolation).
Western blot analysis
The total tissue lysate of the samples from the cortex, striatum, and hippocampus was prepared on ice in cell lysis buffer (150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 1% Triton, 2.5 mM sodiumpyrophosphate, 1 mM β-glycerophosphate, 1 mM Na3VO4, 1 μg/ ml leupeptin; 0.1 ml/100 mg tissue), which was obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA), and contained 2% SDS, protease inhibitor cocktail tablets (Roche Diagnostics Corporation, Indianapolis, IN, USA), and halt protease and phosphatase inhibitor cocktail with EDTA (Thermo Fisher Scientific, Rockford, IL, USA). The samples were then sonicated for 30 seconds using an ultrasonic processor (Sonics and Materials, Inc., Newtown, CT, USA), and centrifuged for 20 min at 4oC and 13000 rpm. The amount of total protein from the supernatant in each sample was quantitatively determined using the Bradford protein assay (Biorad, Hercules, CA, USA). The absorbance was read at 595 nm on a NanoDrop 2000 micro-volume spectrophotometer (Thermo Fisher Scientific, Rockford, IL, USA). Protein concentrations were calculated against a BSA standard curve. For Western blotting, equal amount of protein for each sample was subjected to gradient SDS-PAGE (Biorad; 4-20% Mini-PROTEAN® TGX™ precast polyacrylamide gels; 200 V for 1 h). Protein bands were transferred onto a Immobilon-P PVDF transfer membrane (Millipore, Billerica, MA, USA), blocked for 1 h with 10% non-fat dry milk, washed three times at 10 min each with 1X Tris-Buffered Saline and Tween 20 (TBST), and probed with goat polyclonal primary antibody for dopamine transporter (DAT (C-20), 1:1000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), and rabbit polyclonal primary antibodies for actin (1:5000; Sigma-Aldrich, St. Louis, MO, USA), which was used as a control for sample loading and transfer efficiency, tyrosine hydroxylase (TH, 1:5,000; Millipore, Billerica, MA, USA) and alpha-synuclein antibody (1:2000; Millipore), overnight at 4oC. After four consecutive washes, the membranes were incubated with horseradish peroxidase (HRP) conjugated rabbit anti-goat (1:5000; Abcam, Cambridge, MA, USA) or goat anti-rabbit (1:5000; Promega Corporation, Madison, WI, USA) secondary antibodies for 2 h, washed seven times with 1X TBST, developed using SuperSignal West Pico/Dura/ Femto hemiluminescent substrate (Thermo Fisher Scientific), and exposed to HyBlot CL®autoradiography film (Denville Scientific, Inc., Metuchen, NJ, USA). Densitometry of the protein bands from the scanned autoradiography films was performed using the Image Processing and Analysis in Java Software (ImageJ, National Institutes of Health, Bethesda, MD, USA, http://rsb.info.nih.gov/ij/).The specificity of the alpha-synuclein antibody was confirmed by using Western blotting of purified recombinant α-synuclein protein. Supplemental Figure 1 depicts serial dilution of recombinant alphasynuclein protein (0.01-0.2 μg) that was loaded across the lanes and probed by Western analysis. Supplemental Figure 1B depicts the standard curve generated from the densitometric values obtained from digitized film of the immunoblot. A linear relationship between optical density and the amount of recombinant alpha-synuclein protein exist with a correlation coefficient of 0.95. The lower (~19 kDa) and higher molecular weight species (~37, and >50 kDa) of recombinant human alpha-synuclein peptide are shown.
RNA isolation and qPCR Protocol
Frozen brain structures (striatum, hippocampus, cortex) were homogenized using mortar and pestle in presence of nitrogen. Total RNA was then extracted by means of Trizol (Sigma, USA). RNA was further isolated, cleaned and DNase treated using RNeasy spin columns (Qaigen, USA) according to manufacturers protocol. The RNA quality was assessed by determining 28S and 18S ribosomal RNA integrity using nanoprobe and Bioanalyzer (Bio-rad, USA). Only samples with RIN values of 7 or higher were used in experiments. 1μg of total RNA was then reverse transcribed to cDNA using the iScript cDNA Synthesis Kit (Bio-rad, USA). In short, 5xiScript reaction mix was mixed with reverse transcriptase, nuclease free H2O and RNA template. The mixture was incubated at 5min at 25°C, 30 min at 42°C, 5 min at 85°C and held at 4°C until further use. 20ng of the resulting cDNA was then used to quantitatively measure the expression of genes using SsoFast Evagreen Supermix (Bio-Rad, USA) according to manufacturer’s protocol using the following cycling conditions; activation 30 sec. 95°C, denaturation 5 sec. 95°C and annealing/ extension 5 sec. 60°C. The following primer pairs were used to measure gene expression; alpha-synuclein 5”-TGGGTACCCCTCCTCACCCTT-3” and 5”-GTGGAGCAAAAATACATCTTTAG-3”, Beta-synuclein 5-”CCCGGCCCCGAAACCCAGGCCG -3” and 5”-CTTCAGGTCTGTGGGGAACTCTT-3”,gammasynuclein 5”-CCAGGTCCATCCTTGCAAACA-3” and 5”-CTTTGGCTTCTTGGTTCCTGTG-3” and the housekeeping genes GADPH 5”- CGGCATGTCAGATCCACAAC-3” and 5”-GGCCTTCCGTGTTCCTACC-3”, EF-1 5”-GCAAGCCCATGTGTGTTGAA-3” and 5’-TGATGACACCCAACAGCAACTG-3”. Real time measurements were performed with a Bio-Rad CFX96 Real Time PCR System and analyzed using q-Base plus software (Biogazelle).
Statistical analysis
Differences between groups were determined using GraphPad Prism software by unpaired, two-tailed Student’s t-test. A P-value of <0.05 was considered significant.
Results
Chronic Methamphetamine Exposure Affected Alpha- Synuclein Protein Levels in the Striatum and Hippocampus but Not In the CortexSimilar to our recent report, animals exposed to methamphetamine exhibited increased body temperature and decreased body weight that returned to saline control levels by day 7 [25]. The alpha-synuclein protein level was examined by Western immunoblot analysis in the striatum, hippocampus and cortex (Figure 1B-3B, respectively) of methamphetamine and saline treated animals. Overall, in methamphetamine-exposed animals, we observed a heterogeneous alpha-synuclein accumulation in the striatum, hippocampus and cortex without an effect on the mRNA levels of alpha-, beta- or gamma-synuclein in these regions (Figure 1A-3A).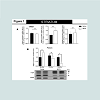 The 19 kDa alpha-synuclein immuno-reactive band was detected in all brain regions examined (Figure 1B-3B). Denser, high molecular weight alpha-synuclein immuno-reactive species (~50 kDa) were detected in the striatum and hippocampus of methamphetamine treated animals with significantly higher optical density as compared to the saline-treated animals (student T-test, p<0.05). The ~50 kDa alpha-synuclein immuno-reactive species were also detected in saline treated animals, but their densities were significantly lower than methamphetamine-treated animals (student T-test, p<0.05).The high molecular weight alpha-synuclein species are thought to represent alpha-synuclein oligomers [26] or ubiquitin modified forms of the protein [27,28]. We did not observe accumulation of these high molecular weight alpha-synuclein species in the cortex of methamphetamine-treated animals when compared to saline treated animals (Figure 2B).
The 19 kDa alpha-synuclein immuno-reactive band was detected in all brain regions examined (Figure 1B-3B). Denser, high molecular weight alpha-synuclein immuno-reactive species (~50 kDa) were detected in the striatum and hippocampus of methamphetamine treated animals with significantly higher optical density as compared to the saline-treated animals (student T-test, p<0.05). The ~50 kDa alpha-synuclein immuno-reactive species were also detected in saline treated animals, but their densities were significantly lower than methamphetamine-treated animals (student T-test, p<0.05).The high molecular weight alpha-synuclein species are thought to represent alpha-synuclein oligomers [26] or ubiquitin modified forms of the protein [27,28]. We did not observe accumulation of these high molecular weight alpha-synuclein species in the cortex of methamphetamine-treated animals when compared to saline treated animals (Figure 2B).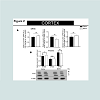 There was a significant decrease in the full-length, 19 kDa, alpha-synuclein protein levels in the striatal region (Figure 1B) (student T-test, p<0.05), but not in the hippocampus or cortical regions of methamphetamine exposed animals as compared to saline treated animals (Figure 2b and 3B). The level of alpha-, beta- and gamma-synuclein mRNA in the striatum, hippocampus and cortex of methamphetamine-exposed animals were similar and were at saline control levels for each brain structure examined (Figure 1A-3A). These results are consistent with the interpretation that the increased striatal and hippocampal alpha-synuclein levels might be consequence of alteration in the translation efficiency of the protein, its stability or its degradation mechanism/s.
There was a significant decrease in the full-length, 19 kDa, alpha-synuclein protein levels in the striatal region (Figure 1B) (student T-test, p<0.05), but not in the hippocampus or cortical regions of methamphetamine exposed animals as compared to saline treated animals (Figure 2b and 3B). The level of alpha-, beta- and gamma-synuclein mRNA in the striatum, hippocampus and cortex of methamphetamine-exposed animals were similar and were at saline control levels for each brain structure examined (Figure 1A-3A). These results are consistent with the interpretation that the increased striatal and hippocampal alpha-synuclein levels might be consequence of alteration in the translation efficiency of the protein, its stability or its degradation mechanism/s.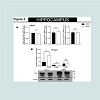 The effects of chronic methamphetamine exposure on two previously reported indicators of methamphetamineinduced neurotoxicity
The effects of chronic methamphetamine exposure on two previously reported indicators of methamphetamineinduced neurotoxicity
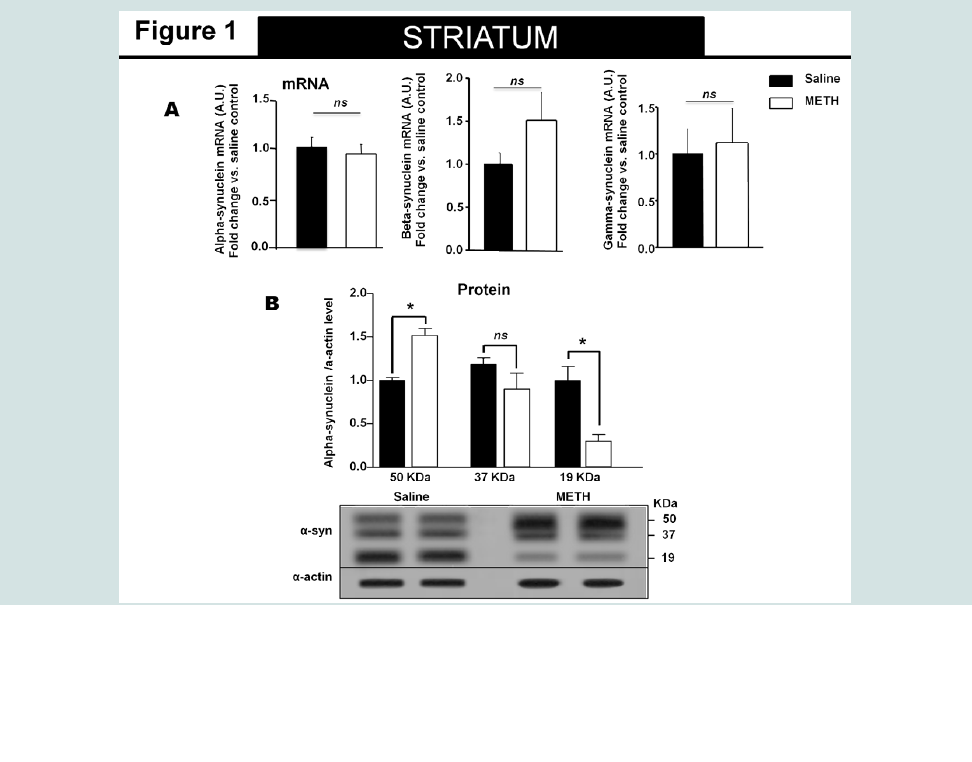
Figure 1: High molecular species of alpha-synuclein protein detected in the striatum of animals exposed to non-contingent methamphetamine for 14 days. The treatment did not change alpha, beta or gamma-synuclein mRNA level in the striatum.
A). Fourteen days of methamphetamine-exposure does not alter striatal mRNA level of alpha-, beta- or gamma-synuclein. Total RNA was obtained from 5–6 animals per group, the mRNA expression was measured individually and normalized to GAPDH and EF1 mRNA levels.
B). Western blot data suggest that fourteen days of methamphetamine treatment increased the relative amount of high molecular species (>50kDa) of alpha-synuclein and decreases the relative amount of low molecular weight (19kDa) of the protein. Equivalent protein samples were loaded. α-actin was used to verify equal protein loading and transfer. The histograms represent the normalized levels of alpha-synuclein expressed as a ratio. The values represent means ± SEM in comparison to the saline-treated group. N=5-6 animals per group. Key to statistics: *, p<0.05.
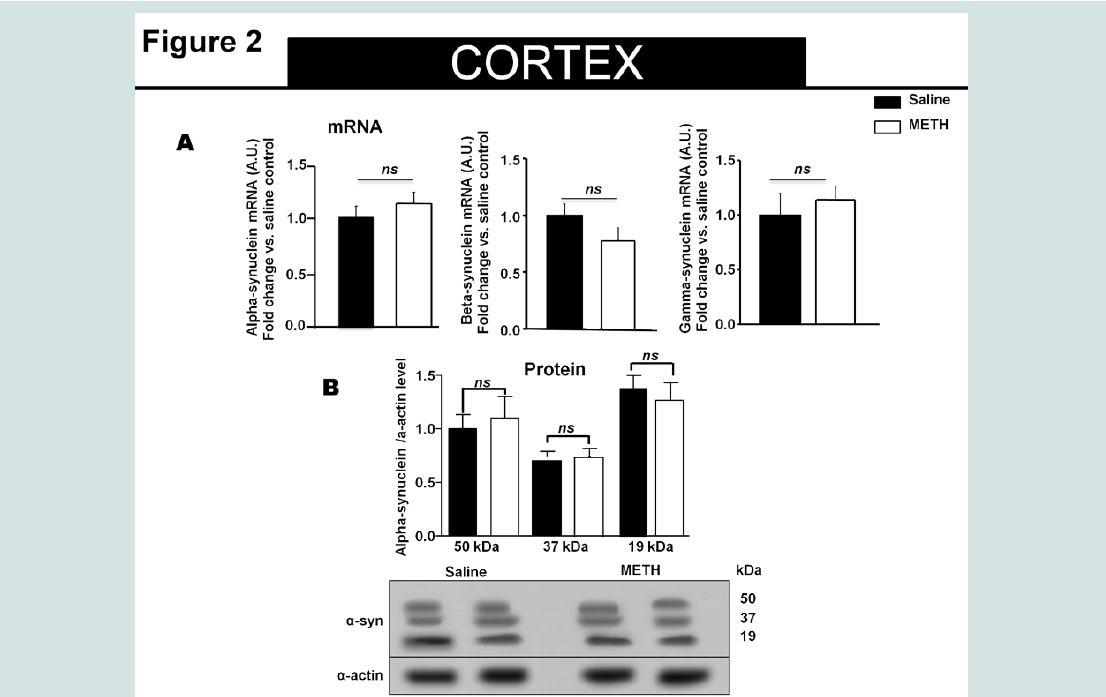
Figure 2: Fourteen days of methamphetamine exposure do not alter the relative levels of alpha-synuclein mRNA and protein in the cortex.
A). Fourteen days of methamphetamine-exposure does not alter cortical mRNA level of alpha-synuclein, beta- and gamma-synuclein. Total RNA was obtained from 5–6 animals per group, the mRNA expression was measured individually and normalized to GAPDH and EF1 mRNA levels.
B). Western blot data suggest that fourteen days of methamphetamine treatment do not change the relative amount of alpha-synuclein levels as compared to saline control. Equivalent protein samples were loaded. α-actin was used to verify equal protein loading and transfer. The histograms represent the normalized levels of alpha-synuclein expressed as a ratio. The values represent means ± SEM in comparison to the saline-treated group. N=5-6 animals per group.
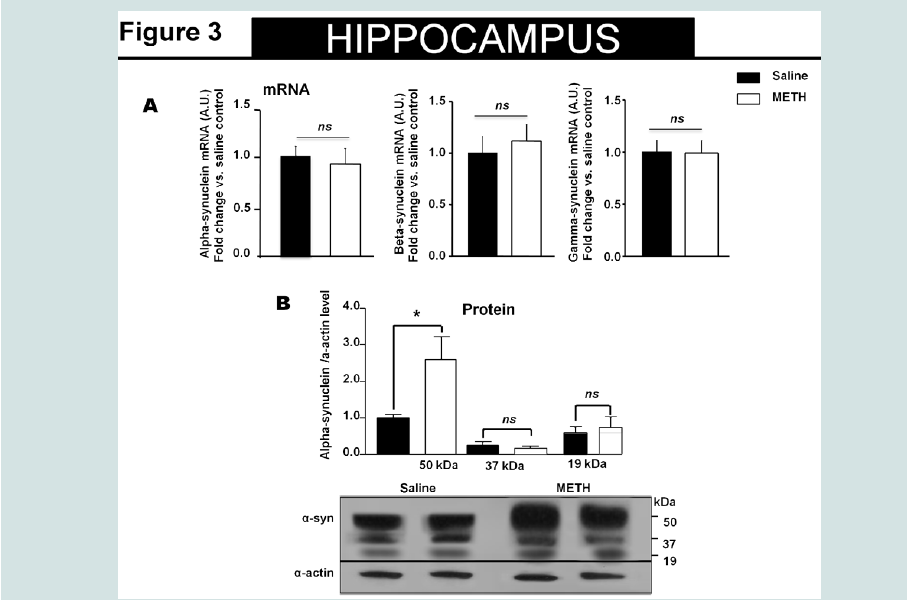
Figure 3: High molecular species of alpha-synuclein protein detected in the hippocampus of animals exposed to non-contingent methamphetaminefor 14 days. The treatment did not change alpha-synuclein mRNA level in the hippocampus.
A). Fourteen days of methamphetamine-exposure does not alter hippocampal mRNA level of alpha-synuclein or beta- and gamma-synuclein. Total RNA was obtained from 5–6 animals per group. The mRNA expression was measured individually and normalized to GAPDH and EF1 mRNA levels.
B). Western blot data suggest that fourteen days of methamphetamine treatment increased the relative amount of high molecular species (>50kDa) of alpha-synuclein without altering the relative amount of low molecular weight (19kDa) of the protein. Equivalent protein samples were loaded. α-actin was used to verify equal protein loading and transfer. The histograms represent the normalized levels of alpha-synuclein expressed as a ratio. The values represent means ± SEM in comparison to the saline-treated group. N=5-6 animals per group. Key to statistics:*, p<0.05.
Consistent with previous reports [25] two weeks of noncontingent neurotoxic methamphetamine exposure produced signs of neurotoxicity as shown by decrease in the tyrosine hydroxylase (TH) (data not shown) and dopamine transporter (DAT) (data not shown) protein levels in the cortex, hippocampus, and striatum of methamphetamine-exposed animals as compared to controls (student T-test, p<0.05, saline compared to methamphetamine exposed animals).
Discussion
One of the major findings of this short report is that multiple immuno-reactive bands for alpha-synuclein were detected in the striatum, hippocampus and cortex of saline treated juvenile mice and that two weeks of methamphetamine exposure altered alphasynuclein protein levels in the striatum and hippocampus but not in the cortex of juvenile mice. Methamphetamine exposure decreased the monomeric form of alpha-synuclein (molecular species <19 kDa)in the striatum while it promoted accumulation of higher molecular weight (>19 kDa) alpha-synuclein species in the striatum and hippocampus but not in the cortex. In contrast to the influence of methamphetamine on alpha-synuclein protein levels, there was no change in the alpha, beta or gamma-synuclein mRNA levels in the striatum, hippocampus and cortex.Consistent with the literature [29] in saline treated mice, various alpha-synuclein immunoreactive bands were detected. Since, it has been reported the nerve cells survive for decades in the presence of immunoreactive alpha-synuclein aggregates [30], it is not known whether alpha-synuclein aggregates are harmless, neuroprotective [31], or harmful to neuronal function [32].
In the striatum of methamphetamine-exposed animals, while the 50kDa molecular species of alpha-synuclein increased, the 19 kDa molecular species are decreased. Although the functional consequences of these biochemical changes are unknown, our data and the literature support the idea that a positive correlation exist between psychostimulant exposure and increased striatal levels of alpha-synuclein. For instance, studies from Mash’s laboratory demonstrated cocaine abusers have an overexpression of alphasynuclein in their dopamine neurons in the ventral tegmental area [12]. In addition, Fornai’s group, reported an increase in the occurrence of alpha-synuclein neuronal inclusions within nigral dopamine neurons of mice following methamphetamine treatment [33]. Therefore, although a positive correlation exists between methamphetamineinduced neurotoxicity and changes in alpha-synuclein protein levels in different brain structures, the causal mechanism is unknown.
While there were increases in alpha-synuclein protein level in the striatum and hippocampus of methamphetamine-treated animals; there was no change in the mRNA level of this protein in the brain structures examined. This is in accordance with the interpretation that methamphetamine may influence the efficiency of alphasynuclein synthesis, its stability, or degradation rate. This possibility is supported by a recent report showing binge methamphetamine exposure alters protein degradation via the ubiquitin proteasomal system leading to dysregulation of protein turnover, accumulation, and potential formation of protein aggregates [34,35]. Nanopore analysis suggests that methamphetamine binds to the N-terminus of α-synuclein and causes a conformational change in the protein [16].Therefore, one of the possible mechanisms for methamphetamine induced elevation of alpha synuclein protein levels can be due to its effect on the degradation of intracellular proteins [36,37]. Overall the mechanism(s) by which methamphetamine increases alphasynuclein protein levels and its direct implications on dopaminergic signaling remains to be examined. Future studies will determine whether the methamphetamine-induced increase in alpha-synuclein protein levels is one of the causal mechanisms for methamphetaminemediated neurotoxicity, or it is one of the compensatory protective mechanisms.
Acknowledgement
Funding sources for this manuscript DA026947/DA/NIDA NIH HHS/ United States and NS071122/NS/NINDS NIH HHS/United States.References
- Vearrier D, Greenberg MI, Miller SN, Okaneku JT, Haggerty DA (2012) Methamphetamine: history, pathophysiology, adverse health effects, current trends, and hazards associated with the clandestine manufacture of methamphetamine. Dis Mon 58: 38-89.
- Logan BK (1996) Methamphetamine and driving impairment. J Forensic Sci 41: 457-464.
- Bell DS (1973) The experimental reproduction of amphetamine psychosis. Arch Gen Psychiatry 29: 35-40.
- Wilson JM1, Kalasinsky KS, Levey AI, Bergeron C, Reiber G, et al. (1996) Striatal dopamine nerve terminal markers in human, chronic methamphetamine users. Nat Med 2: 699-703.
- Cadet JL, Jayanthi S, Deng X (2003) Speed kills: cellular and molecular bases of methamphetamine-induced nerve terminal degeneration and neuronal apoptosis. FASEB J 17: 1775-188.
- Kita T, Wagner GC, Nakashima T (2003) Current research on methamphetamine-induced neurotoxicity: animal models of monoamine disruption. J Pharmacol Sci 92: 178-195.
- Hashimoto K1, Tsukada H, Nishiyama S, Fukumoto D, Kakiuchi T, et al. (2004) Protective effects of N-acetyl-L-cysteine on the reduction of dopamine transporters in the striatum of monkeys treated with methamphetamine. Neuropsychopharmacology 29: 2018-2023.
- Zhang L, Kitaichi K, Fujimoto Y, Nakayama H, Shimizu E, et al. (2006) Protective effects of minocycline on behavioral changes and neurotoxicity in mice after administration of methamphetamine. Prog Neuropsychopharmacol Biol Psychiatry 30: 1381-1393.
- Baucum AJ , Rau KS, Riddle EL, Hanson GR, Fleckenstein AE (2004) Methamphetamine increases dopamine transporter higher molecular weight complex formation via a dopamine- and hyperthermia-associated mechanism. J Neurosci 24: 3436-3443.
- Callaghan RC, Cunningham JK, Sykes J, Kish SJ (2012) Increased risk of Parkinson's disease in individuals hospitalized with conditions related to the use of methamphetamine or other amphetamine-type drugs. Drug Alcohol Depend 120: 35-40.
- Callaghan RC, Cunningham JK, Sajeev G, Kish SJ (2010) Incidence of Parkinson's disease among hospital patients with methamphetamine-use disorders. Mov Disord 25: 2333-2339.
- Mash DC, Ouyang Q, Pablo J, Basile M, Izenwasser S, et al. (2003) Cocaine abusers have an overexpression of alpha-synuclein in dopamine neurons. J Neurosci 23: 2564-2571.
- Ma QL, Chan P, Yoshii M, Uéda K, (2005) Alpha-synuclein aggregation and its relation to neurodegenerative diseases. J Alzheimers Dis 57: 605-606.
- Qin Y, Ouyang Q, Pablo J, Mash DC, et al. (2005) Cocaine abuse elevates alpha-synuclein and dopamine transporter levels in the human striatum. Neuroreport 16: 1489-1493.
- Fornai F1, Lenzi P, Ferrucci M, Lazzeri G, di Poggio AB, et al. (2005) Occurrence of neuronal inclusions combined with increased nigral expression of alpha-synuclein within dopaminergic neurons following treatment with amphetamine derivatives in mice. Brain Res Bull 65: 405-413.
- Tavassoly O, Lee JS (2012) Methamphetamine binds to alpha-synuclein and causes a conformational change which can be detected by nanopore analysis. FEBS Lett 586: 3222-3228.
- Spear LP (2004) Adolescent brain development and animal models. Ann N Y Acad Sci 1021: 23-26.
- Spear LP (2000) The adolescent brain and age-related behavioral manifestations. Neurosci Biobehav Rev 24: 417-463.
- Fumagalli F, Gainetdinov RR, Valenzano KJ, Caron MG (1998) Role of dopamine transporter in methamphetamine-induced neurotoxicity: evidence from mice lacking the transporter. J Neurosci 18: 4861-4869.
- Wallace TL, Gudelsky GA, Vorhees CV (1999) Methamphetamine-induced neurotoxicity alters locomotor activity, stereotypic behavior, and stimulated dopamine release in the rat. J Neurosci 19: 9141-9148.
- Armstrong BD, Noguchi KK (2004) The neurotoxic effects of 3,4-methylenedioxymethamphetamine (MDMA) and methamphetamine on serotonin, dopamine, and GABA-ergic terminals: an in-vitro autoradiographic study in rats. Neurotoxicology 25: 905-914.
- Cadet JL, Brannock C, Ladenheim B, McCoy MT, Beauvais G, et al. (2011) Methamphetamine preconditioning causes differential changes in striatal transcriptional responses to large doses of the drug. Dose Response 9: 165-181.
- Sonsalla PK, Jochnowitz ND, Zeevalk GD, Oostveen JA, Hall ED, et al. (1996) Treatment of mice with methamphetamine produces cell loss in the substantia nigra. Brain Res 738: 172-175.
- O'Callaghan JP, Miller DB (1994) Neurotoxicity profiles of substituted amphetamines in the C57BL/6J mouse. J Pharmacol Exp Ther 270: 741-751.
- North A, Swant J, Salvatore MF, Gamble-George J, Prins P, et al. (2013) Chronic methamphetamine exposure produces a delayed, long-lasting memory deficit. Synapse. 67: 245-257.
- Sharon R, Bar-Joseph I, Frosch MP, Walsh DM, Hamilton JA, et al. (2003) The formation of highly soluble oligomers of alpha-synuclein is regulated by fatty acids and enhanced in Parkinson's disease. Neuron 37: 583-595.
- Sampathu DM, Giasson BI, Pawlyk AC, Trojanowski JQ, Lee VM (2003) Ubiquitination of alpha-synuclein is not required for formation of pathological inclusions in alpha-synucleinopathies. Am J Pathol 163: 91-100.
- Apetri MM, Maiti NC, Zagorski MG, Carey PR, Anderson VE (2006) Secondary structure of alpha-synuclein oligomers: characterization by raman and atomic force microscopy. J Mol Biol 355: 63-71.
- Tong J, Wong H, Guttman M, Ang LC, Forno LS, et al. (2010) Brain alpha-synuclein accumulation in multiple system atrophy, Parkinson's disease and progressive supranuclear palsy: a comparative investigation. Brain 133:172-188.
- Goedert M, Spillantini MG, Del Tredici K, Braak H (2013) 100 years of Lewy pathology. Nat Rev Neurol 9: 13-24.
- Saha AR, Hill J, Utton MA, Asuni AA, Ackerley S, et al. (2004) Parkinson's disease alpha-synuclein mutations exhibit defective axonal transport in cultured neurons. J Cell Sci 117: 1017-1024.
- Del Tredici K, Braak H (2012) Lewy pathology and neurodegeneration in premotor Parkinson's disease. Mov Disord 27: 597-607.
- Mauceli G, Busceti CI, Pellegrini A, Soldani P, Lenzi P, et al. (2006) Overexpression of alpha-synuclein following methamphetamine: is it good or bad? Ann N Y Acad Sci 1074:191-197.
- Fornai F1, Lenzi P, Gesi M, Ferrucci M, Lazzeri G, et al. (2004) Similarities between methamphetamine toxicity and proteasome inhibition. Ann N Y Acad Sci 1025: 162-170.
- Moszczynska A, Yamamoto BK (2011) Methamphetamine oxidatively damages parkin and decreases the activity of 26S proteasome in vivo. J Neurochem 116: 1005-1017.
- Staszewski RD, Yamamoto BK (2006) Methamphetamine-induced spectrin proteolysis in the rat striatum. J Neurochem 96: 1267-1276.
- Straiko MM, Coolen LM, Zemlan FP, Gudelsky GA (2007) The effect of amphetamine analogs on cleaved microtubule-associated protein-tau formation in the rat brain. Neuroscience 144: 223-231.