
Review Article
Corresponding author: Jiansheng Liang, Department of Biology, College of Bioscience and Biotechnology, Yangzhou University, 12 East Wenhui Road, Yangzhou, Jiangsu, PR CHINA. 225009, Tel: 086 755 8801 0053; Fax: 086 755 88010035; E-mail: jsliang@yzu.edu.cn
Citation: Dongping Z, Li C, Bing LV, Jiansheng L. The Scaffolding Protein RACK1: A Platform for Diverse Functions in the Plant Kingdom. J Plant Biol Soil Health. 2013;1(1): 7.
Copyright © 2013 Dongping Z, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Biology & Soil Health | ISSN: 2331-8996 | Volume: 1, Issue: 1
Submission: 05 August 2013 | Accepted: 29 August 2013 | Published: 04 September 2013
The Scaffolding Protein RACK1: A Platform for Diverse Functions in the Plant Kingdom
ZHANG Dongping1, CHEN Li1, LV Bing1 and LIANG Jiansheng1,2
- 1College of Bioscience and Biotechnology, Yangzhou University, P. R. China, 225009
- 2Department of Biology, South University of Science and Technology of China, Shenzhen, P. R. China, 518055
Corresponding author: Jiansheng Liang, Department of Biology, College of Bioscience and Biotechnology, Yangzhou University, 12 East Wenhui Road, Yangzhou, Jiangsu, PR CHINA. 225009, Tel: 086 755 8801 0053; Fax: 086 755 88010035; E-mail: jsliang@yzu.edu.cn
Citation: Dongping Z, Li C, Bing LV, Jiansheng L. The Scaffolding Protein RACK1: A Platform for Diverse Functions in the Plant Kingdom. J Plant Biol Soil Health. 2013;1(1): 7.
Copyright © 2013 Dongping Z, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Biology & Soil Health | ISSN: 2331-8996 | Volume: 1, Issue: 1
Submission: 05 August 2013 | Accepted: 29 August 2013 | Published: 04 September 2013
Abstract
The receptor for activated C kinase 1 (RACK1) is one of the well-known WD-repeat containing proteins and can interact as a scaffolding protein with many different signaling proteins from distinct signal transduction pathways. Accordingly, it acts as a platform for diverse functions in both prokaryotes and eukaryotes. In metazoan and yeast, RACK1 is known to be involved in regulating development, circadian rhythm, cell migration, diseases, etc. However, the roles of RACK1 are largely unclear in plant kingdom. Nevertheless, great progresses have been made in recent years on the roles of plant RACK1. Plant RACK1 is highly similar to mammalian RACK1 at the amino acid level and in structure, although some differences in amino acid sequence of RACK1 proteins have been found between monocots and dicots as well as between human and plants, which can explain, at least in part, the differences of RACK1 functions. Several recent studies have shown that plant RACK1 is involved in the responses to plant hormones, abiotic/biotic stresses, plant growth and development, etc. And the molecular mechanisms of RACK1 functions have also been explored, although the gap is still obvious between plants and metazoan. However, a deep understanding of the molecular mechanisms of RACK1 functions is expected as the progress in screening and identification of more and more RACK1-interacting signaling molecules.Keywords
Heterotrimic G protein; Plant growth and development; RACK1; Scaffolding protein; WD-repeatIntroduction
The receptor for activated C kinase 1 (RACK1) is one member of the most important tryptophan, aspartic acid repeat (WD repeat)- containing family of proteins and has been extensively studied during past two decades, especially in metazoan and yeast [1-6]. Now it is wildly recognized that RACK1 protein is strongly conserved from prokaryotes to eukaryotes and expresses ubiquitously [7,8]. The detailed structure characterization of RACK1 has been clarified recently in several organisms at different levels of solution, which shares significant homology to the β subunit of the heterotrimeric G protein (G protein) and provides structural basis to shed light on the molecular mechanisms of RACK1 in the regulation of diverse signaling pathways as a scaffolding protein [5,6,9,10]. Compared to the advance obtained from the studies in metazoan and yeast, we know little about the functions and molecular mechanisms of plant RACK1. In this review, we focus on the recent progresses on plant RACK1. Readers can also refer to several excellent review articles and the references therein [7,8].RACK1 is a Highly Conserved WD-40 Protein Found in Both Prokaryotes and Eukaryotes
RACK1, originally identified as a receptor for activated protein kinase C (PKC), belongs to one of the WD-repeat-containing protein family found in prokaryotes and all eukaryotes, and is involved in multiple signaling pathways [11-16]. The first RACK1 gene was cloned from a chicken liver cDNA library [17], and since then RACK1 gene was isolated from a range of species, including Arabidopsis thaliana, Oryza sativa, Tetrahymena thermophile, Homo sapiens, Rattus norvegicus, Caenorhabditis elegans, Drosophila melanogaster, green algae, Saccharomyces cerevisiae, Phaseolus vulgaris, Nicotiana tabacum, Medicago sativa, etc. [18-29]. Sequence alignments of RACK1 species from diverse organisms reveal a 43-76% sequence identity at the amino acid level, indicating that RACK1 is strongly conserved through evolution [7,8].
Structurally, RACK1 protein contains seven WD domains, which itself is sequences of typically 44-60 amino acids in length and starts with a glycine-histidine (GH) dipeptide 11 to 24 residues from the N terminus and end with a signature tryptophan-aspartic acid (WD) dipeptide at the C terminus [27], though neither the GH nor WD signature is absolutely conserved. The high sequence identity and similar repeating segments have led to the prediction that RACK1 proteins from different organisms share a common or at least similar structural characterization, which is verified by the resolution of the crystal structures of RACK1 proteins of several organisms fromdifferent laboratories, including those from Saccharomyces cerevisiae [5], Arabidopsis thaliana [10] and Tetrahymena thermophila [6], Homo sapiens [9] at 2.1, 2.4, 3.9 and 2.45Å resolution, respectively. These structural studies confirm that RACK1 adopts a seven-bladed β -propeller structure consistent with the structure of G-protein β subunit (Gβ), which is the first WD repeat protein structurally characterized by X-ray crystallography [28,29]. The strong similarity in structure of RACK1 from different organisms is in accordance with the high conservation of the amino acid sequences. And also this structural characterization suggests that the availability of multiple protein interaction surfaces like many other WD repeat containing proteins, thereby serving as hubs for spatiotemporal orchestration of signaling events across diverse pathways.
The Structural Bases of RACK1 as a Scaffolding Protein
As described above, RACK1 belongs to the WD-40 repeat family which is a highly conserved intracellular adaptor protein with significant homology to Gβ. Gβ was the first structurally characterized WD domain-containing protein, which adopts a seven-bladed β-propeller structure and exists in a complex with the γ subunit (Gγ). X-ray crystallographic study shows that Gβ structure is composed of seven blades, each containing a four-stranded anti-parallel β-sheet, connected by loops. These compact and multisided structural features allow Gβγ to bind reversibly with α subunit to form a Gαβγ heterotrimer. It is now well established that Gβ, as a platform for scaffolding protein, interacts with a large number of signal molecules and plays crucial roles in various signaling cascades. Similar structural feature of RACK1 to Gβ has been characterized in different organisms, which all contain seven WD-repeat domains and fold into seven β-propeller tertiary structure [5,7,10,30]. Like many other WD proteins, there exist multiple protein-binding sites within RACK1 molecule, which enables it interact with a large number of proteins either directly or as part of a larger complex and integrate inputs from distinct signaling pathways. This scaffolding property of RACK1 is mediated by the presence of seven WD-repeats and any change in WD-repeats may significantly affect the interaction with its binding partners, such as its specificity/selectivity and/or binding properties. In fact, despite of the strongly conserved amino acid sequences of RACK1 proteins in different organisms, the available information about RACK1 crystal structure indicates that this conservation is not absolute and some variations exist in both the composition and the numbers of amino acids (Figure 1). Great variations in the length of GH-WD core sequences are observed between the blade 1 and 2, blade 3 and 4, and blade 6 and 7 in the RACK1 species, which seriously affects the final tertiary structure of RACK1, and as the consequence, the interaction with its partners. For example, the greatest variable region between blades 6 and 7 is predicted to form a pronounced knob-like projection from the upper face of the propeller, thus presumably determining their binding properties. For plant RACK1, great variations are also found between monocots and dicots, of which the differences are mainly located in WD-domain 1, 3 and 7 (Figure 1). We now know little about whether it means that there are some differences in structure and functions of RACK1 between monocots and dicots. In other words, the multiple protein-binding sites presented in RACK1 protein and the variations in RACK1 structure provide the structural bases as the scaffold proteins that facilitate interaction with specialized protein docking modules and that explain the specialized roles played in different organisms. The efficiency and specificity of signal transduction are largely determined by the protein-protein interaction, which is very similar to an enzymatic catalyzing reaction within the cell. Scaffolding proteins are located at the central position of the cell-signaling network and contribute to the accuracy of the spatiotemporal processing and transmission of a signal. As a scaffold protein, RACK1 protein assembles signaling components of a cascade from different signaling pathways into complexes. This assembly may be able to enhance signaling specificity and efficiency by preventing unnecessary interactions between signaling proteins and by increasing the proximity and effective concentration of components in the scaffold complex, respectively [7]. Generally, RACK1 interacts with a large number of proteins either directly or as part of a larger complex either as a monomer form or as a homodimer. The later enables RACK1 to bind more signaling molecules and modulate more cellular functions. Understandably, the molecular mechanisms of RACK1 functions are also largely dependent upon the cell types as well as the state of the cells.
The efficiency and specificity of signal transduction are largely determined by the protein-protein interaction, which is very similar to an enzymatic catalyzing reaction within the cell. Scaffolding proteins are located at the central position of the cell-signaling network and contribute to the accuracy of the spatiotemporal processing and transmission of a signal. As a scaffold protein, RACK1 protein assembles signaling components of a cascade from different signaling pathways into complexes. This assembly may be able to enhance signaling specificity and efficiency by preventing unnecessary interactions between signaling proteins and by increasing the proximity and effective concentration of components in the scaffold complex, respectively [7]. Generally, RACK1 interacts with a large number of proteins either directly or as part of a larger complex either as a monomer form or as a homodimer. The later enables RACK1 to bind more signaling molecules and modulate more cellular functions. Understandably, the molecular mechanisms of RACK1 functions are also largely dependent upon the cell types as well as the state of the cells.
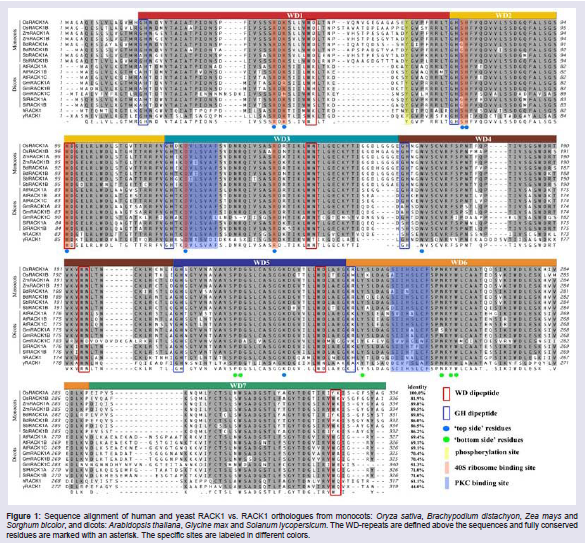
Figure 1: Sequence alignment of human and yeast RACK1 vs. RACK1 orthologues from monocots: Oryza sativa, Brachypodium distachyon, Zea mays and Sorghum bicolor, and dicots: Arabidopsis thaliana, Glycine max and Solanum lycopersicum. The WD-repeats are defined above the sequences and fully conserved residues are marked with an asterisk. The specific sites are labeled in different colors.
It is well known that RACK1 itself does not have the enzymatic activity, but it may possess a catalytic-like property as interaction with signaling proteins may result in allosteric changes of these signaling components. Such changes may be able to promote or suppress the activation of these signaling proteins [31-34]. When cells are triggered by an external stimulus, the RACK1 protein interacts with its signaling partners and activates or inactivates the latters, as a consequence, transducing the external signal into intracellular signal and resulting in the changes of cellular processes. In some cases, RACK1 acts as a transporter to shuttle its binding partners to the particular intracellular sites and enable its partners to exert their regulatory roles.
Taken together, RACK1 as a scaffolding protein exerts its regulatory roles on a wide range of discrete signaling events with different molecular mechanisms and at the level of several subcellular compartments by controlling the interaction network with different families of signaling proteins. And also, RACK1 functions may vary with the cell types as well as the state of the cells.
RACK1 Functions in Plant Kingdom
RACK1 and Plant Hormone Responses and DevelopmentIn past two decades, plenty of results have shown that RACK1 was expressed ubiquitously in different tissues and organs, including leaf, stem, root, and flower, etc., which implies that RACK1 may play important roles in plant growth and development and the responses of plants to biotic and abiotic stresses. Because RACK1 was originally identified as an auxin inducible gene, arcA, in tobacco BY-2 suspension cells in a differential screen for genes involved in auxin-mediated cell division [18,19], it is reasonably to propose that RACK1 are involves in the plant hormone-mediated molecular and physiological processes. Chen et al. provided direct genetic evidence of the function of RACK1 in plant responses to several plant hormones using the loss-of-function mutants of RACK1 in Arabidopsis. They found that rack1a mutants displayed altered sensitivities to several plant hormones, including hyposensitivity to gibberellic acid and brassinosteroid in seed germination, hyposensitivity to auxin in adventitious and lateral root formation, and hypersensitivity to abscisic acid in seed germination and early seedling development [35]. Further study showed that three Arabidopsis RACK1 genes act redundantly to regulate ABA responses in seed germination, cotyledon greening and root growth, because rack1a single and double mutants are hypersensitive to ABA in each of these processes, whereas RACK1A-overexpressed plants displayed ABA insensitivity. The ABA-responsive marker genes, RD29B and RAB18, were upregulated in rack1a mutants and the expression of all three RACK1 genes themselves was down-regulated by ABA [36]. Islas-Flores et al., using a RNAi approach to suppress the RACK1 gene expression in P.vulgaris (PvRACK1), found that mRNA accumulation of PvRACK1 in roots was induced by auxins, abscisic acid, cytokinin, and gibberellic acid [37]. Our studies on the functions of Arabidopsis and rice RACK1 in the responses to drought stress showed that, when RACK1 gene expression was suppressed, leaf ABA level increased significantly, as a result, the tolerance of seedlings to soil drying increased [38].
In addition, growing evidence has shown that RACK1 is a critical regulator of plant development and loss-of-function mutations in RACK1A confer defects in multiple developmental processes including seed germination, leaf production, and flowering. Guo et al. reported that rack1a mutants displayed reduced sensitivity to gibberellin and brassinosteroid in seed germination, hypersensitivity to abscisic acid in seed germination and early seedling development, and hyposensitivity to auxin in adventitious and lateral root formation 35]. Further study has showed that RACK1 genes function in an unequally redundant manner. Both the difference in RACK1 gene expression level and the cross-regulation are likely the molecular determinants of their unequal genetic redundancy [39]. Recently, Islas-Flores et al. studied, using RNA interference technique to silence the PvRACK1 expression in hairy roots of P. vulgaris, the roles of RACK1 in nodulation and development. When the expression of PvRACK1 gene was suppressed, the numbers of nodules formed were 70-90% less than those of non-transgenic roots and the formed nodules were also much smaller. The nodulation inhibition in RACK1-suppressed roots is largely due to the effects of changes of endogenous plant hormones. Whereas the small sizes of nodules are largely due to the inhibition of cell expansion within them [40]. Taken together, RACK1 may be an integrator of diverse signal-transduction pathways in the regulation of plant growth and development.
RACK1 and the Responses to Abiotic and Biotic Stresses
Plants are frequently exposed to various abiotic and biotic stresses, such as drought, salinity, extreme temperature and pathogens, etc., as a result, their growth and development are seriously affected [41]. However, plants have evolved several physiological and molecular mechanisms to cope with these stresses, although we knew little about the detailed of these mechanisms. Direct and indirect evidence have shown that RACK1 are involved in the regulation of plant tolerance to abiotic and biotic stresses [42]. We found the rack1 mutant is strongly tolerant to soil drying as compared with the wild type, Col (Figure 2). Further study indicated that the water loss of detached leaves and stomatal conductance of rack1 mutant are significantly lower than those of wildtype, whereas, the endogenous ABA content of rack1 mutant is higher than that of wild type [39]. Similar results were observed in rice plant, where the RACK1-over-expressed transgenic rice lines were more sensitive to soil drying, whereas the RACK1-suppressed transgenic rice lines more tolerant to soil drying [44]. Comparative proteomic analysis has shown that RACK1 may play roles in regulating plant resistance to salt and heat stresses [40,43]. Recently, Kundu et al. used Arabidopsis RACK1A as a bait to screen a split-ubiquitin based cDNA library and found that about 25 of RACK1A-interacting proteins were directly or indirectly involved with environmental stress signaling pathways [42]. In addition to the involvement of RACK1 in the regulation of plant responses to abiotic stresses, RACK1 was also reported to function in plant innate immunity. Early study showed that RACK1A is involved in control of ROS production, defense gene expression, and disease resistance; all of which are regulated by GTP-bound form of Rac1, small GTPase [44]. In rice, the innate immunity was regulated by a complex of regulatory proteins at the plasma membrane that was composed of RACK1, Rac1, RAR1, SGT1 and two HSP proteins, HSP90 and HSP70. Rac1 transcriptionally and post-transcriptionally regulates RACK1 and vice versa. RACK1A could, as a scaffolding protein, also bind the N terminus of RbohB protein (NADPH oxidase) and play key roles to help assembly and maintain the integrity of this immune complex. When rice plant was attacked by pathogen (i.e. rice blast fungus), Rac1 and/or RACK1 was activated and the immune complex formed. The formed immune complex either directly regulates the immune response or interacts with the N terminus of Rboh to trigger the production of ROS and the disease resistance [45]. Recently, Wamaitha et al. reported that OsRap2.6, a transcription factor, also contributes to rice innate immunity through its interaction with RACK1A [46].
In addition to the involvement of RACK1 in the regulation of plant responses to abiotic stresses, RACK1 was also reported to function in plant innate immunity. Early study showed that RACK1A is involved in control of ROS production, defense gene expression, and disease resistance; all of which are regulated by GTP-bound form of Rac1, small GTPase [44]. In rice, the innate immunity was regulated by a complex of regulatory proteins at the plasma membrane that was composed of RACK1, Rac1, RAR1, SGT1 and two HSP proteins, HSP90 and HSP70. Rac1 transcriptionally and post-transcriptionally regulates RACK1 and vice versa. RACK1A could, as a scaffolding protein, also bind the N terminus of RbohB protein (NADPH oxidase) and play key roles to help assembly and maintain the integrity of this immune complex. When rice plant was attacked by pathogen (i.e. rice blast fungus), Rac1 and/or RACK1 was activated and the immune complex formed. The formed immune complex either directly regulates the immune response or interacts with the N terminus of Rboh to trigger the production of ROS and the disease resistance [45]. Recently, Wamaitha et al. reported that OsRap2.6, a transcription factor, also contributes to rice innate immunity through its interaction with RACK1A [46].
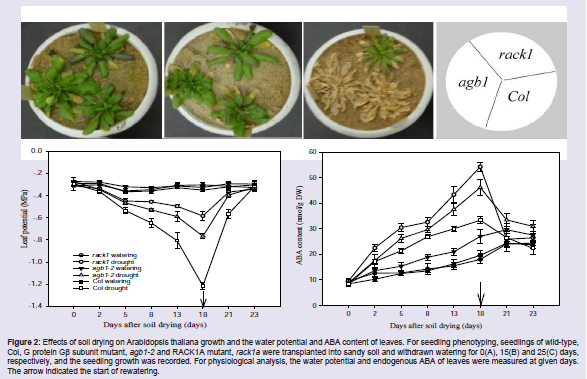
Figure 2: Effects of soil drying on Arabidopsis thaliana growth and the water potential and ABA content of leaves. For seedling phenotyping, seedlings of wild-type,Col, G protein Gβ subunit mutant, agb1-2 and RACK1A mutant, rack1a were transplanted into sandy soil and withdrawn watering for 0(A), 15(B) and 25(C) days, respectively, and the seedling growth was recorded. For physiological analysis, the water potential and endogenous ABA of leaves were measured at given days. The arrow indicated the start of rewatering.
RACK1 and Protein Synthesis
Protein synthesis is a highly regulated process and involves many steps and different subcellular locations. The last step of protein synthesis occurs in the ribosomes, protein synthesis machinery, and so that the composition and abundance of ribosomes are largely related to the efficiency and specificity of protein synthesis. However, several recent studies have shown that ribosome acts not only as the protein synthesis machinery, but as a signaling platform that has essential functions in cellular signaling pathways. First evidence for this comes from the studies on the ribosome structure in human and yeast using cryoelectron microscopy (Cryo-EM) techniques, in which RACK1 and its orthologous have been found to be associated with the 40S small ribosomal subunit [30,47]. Thereafter, Chang et al. found that Arabidopsis orthologous of RACK1 associate with the 40S subunit of cytosolic ribosomes [48]. The same result was also found in algae [49]. The molecular basis of the binding of RACK1 to 40S ribosome subunit has been explored in detailed, but it is largely unknown in plants. Furthermore, Arabidopsis RACK1 may also associate with the 80S ribosomes because both can co-migrate during gradient centrifugation [50]. A recent study using proteomic technique has shown that sucrose feeding increases the abundance of RACK1 protein family members on Arabidopsis ribosome [51]. In another independent study, Guo et al. found that more than 80% of the genes coexpressed with RACK1 encode ribosome proteins and RACK1 physically interacts with Arabidopsis eukaryotic initiation factor 6 (eIF6), whose mammalian homolog is a key regulator of 80S ribosome assembly [52]. These results suggest that RACK1 may be required for normal production of 60S and 80S ribosomes. These results implied that RACK1 had a dual role in signaling and protein synthesis regulation as the cases recognized in mammals and yeasts [47, 53-56].
RACK1 and Heterotrimeric G Protein-Mediated Signaling
Both heterotrimeric GTP-binding proteins (G proteins) and RACK1 play key roles in the regulation of many diverse signal transduction pathways. The high structure similarity between G protein β subunit (Gβ) and RACK1 led to suppose they have some functional relations and/or share similar mechanisms. In other word, they may share the same signal transduction pathway or constitute a signaling complex to regulate these diverse responses. In mammals, RACK1 physically interact not only with the Gβ subunit to form a heterodimer, but with the Gβγ dimer to form heterotrimer [57-59], both of which enable efficient cross-talk between signal transduction pathways mediated by G-protein and RACK1. In yeast, RACK1 (Asc1) can interact with Gβ and Gα and negatively regulate the glucose response mediated by G protein [61]. However, it is largely unclear whether RACK1 is involved in G-protein-mediated signaling processes, or vice versa, in higher plants. Komatsu et al. used a proteomic approach to analyze protein expression profiles in the embryos of rice d1 mutant, a loss of function mutant of the heterotrimeric G protein α subunit (Gα). RACK1 was found to be one of the seven proteins whose expression is down-regulated in d1 mutant. RACK1 protein was induced by abscisic acid in imbibed seeds of wild-type, but not in the d1 mutant [61]. Similar result has also been obtained in Arabidopsis, where the RACK1 gene expression was significantly suppressed in gpa1 mutant. However, these results do not indicate that there exists direct relationship between RACK1 and G protein, because no interactions between RACK1 and Gα/Gβ have been identified either in a yeast split-ubiquitin assay or in Co-IP assay [38,39]. More recent study has shown that RACK1A can interact in vitro with two heterotrimeric G protein gamma subunits, AGG1 and AGG2, and regulate cellular responses to diverse biotic and abiotic stresses [61]. Taken together, the molecular mechanisms of RACK1- and G protein-mediated signaling are very different between plants and mammals.
Rack1 Interacting Partners in Plants
By controlling the interaction network between these different families of signaling proteins, RACK1 regulates a wide range of discrete signaling events. The special structural characterization of RACK1 enables it act as a scaffolding protein to form preassembled or signal dependent multi-protein complexes that contribute to the accuracy of the spatiotemporal processing and transmission of a signal. Therefore, it is the key step to identify RACK1 interacting partners in order to explore the cell-signaling network mediated by RACK1. There are many methods to be used to screen the protein interacting partners. In metazoan, about 90 different proteins have been identified to interact with RACK1 [7]. Recently, Kundu et al. use Arabidopsis RACK1A as a bait to screen a split-ubiquitin based cDNA library and find that 97 proteins potentially interact with RACK1A. These proteins are supposed to involve in plant responses to dehydration and salt stress, protein biosynthesis and photosynthesis pathways [40]. We use the yeast split-ubiquitin system combined with pull-down technique to screen the potential proteins that interact with RACK1 in rice and totally 112 proteins have been identified. These proteins are involved in plant responses to biotic and abiotic stresses, photosynthesis, ROS production, secondary metabolism, protein synthesis, etc. All these provide the basis to shed light on the possible molecular mechanisms of RACK1 in the regulation of different intracellular signal transduction pathways.
Summary and Perspectives
As compared with the progresses in the field of mammalian RACK1, our understanding of the functions and molecular mechanisms of RACK1 in plant is still in its infancy. Although RACK1 is strongly conserved both in amino acid sequence domains and in structure from prokaryotes to eukaryotes, some differences have also been detected among different organisms, which can explain, at least in part, the differences in RACK1 functions. For example, mammalian RACK1 can interact with the Gβ subunit which implies involvement of RACK1 in G protein-mediated signaling in cells. However, no interaction is detected between RACK1 and Gβ subunit in plants so far, although it does not mean that RACK1 has no relation to G protein-mediated signaling in plant cells [61]. Even for plants, there are still some differences in the amino acid sequence of RACK1 protein between monocots and dicots (Figure 1). In addition, unlike the cases in mammalian and yeast, most plant species have more than one copy RACK1 genes, which may make it more difficult to study the RACK1 functions. Therefore it is urgently needed to explore the potential functions of RACK1 in various kinds of plants and clarify if the differences in the amino acid sequence of RACK1 and/or its structure can explain the differences of functions among different plant species, and if there is a potential functional redundancy among these RACK1 genes within the same plant cells.Furthermore, at present, we know little whether plant RACK1 functions as a scaffolding protein like the case in mammalian. If RACK1 indeed acts as a scaffolding protein in plant cells, then it is expected that there are a wide range of interacting proteins with RACK1. However, only small numbers of proteins have been characterized to physically interact with RACK1 in plants. Therefore, one of the future studies may focus on the identification and characterization of the competent RACK1-binding partners in plants, because it is critical to unravel the molecular mechanisms of RACK1 action.
Finally, several other aspects about RACK1 are also of importance in the illumination of the molecular mechanisms of RACK1 functions. For example, whether the post-translational modification of RACK1 is related to its function and whether the plant cell types and physiological state can affect its functions. And also, whether external environmental cues and plant hormones can affect the binding properties between RACK1 and its partners, and the expression patterns and distribution of RACK1 within the plant cells, as a consequence, affect the plant growth and development.
Acknowledgements
This research was supported by grant from the Natural Science Foundation of China (grant no: 31271622) and the Hi-Tech Research and Development Program (“863” project) from the Ministry of Science and Technology of China (grant no: 2008AA10Z120) to JSL.References
- Ron D, Chen CH, Caldwell J, Jamieson L, Orr E, et al. (1994) Cloning of an intracellular receptor for protein kinase C: a homolog of the beta subunit of G proteins. Proc Natl Acad Sci USA 91: 839-43.
- Xu C, Min J(2011) Structure and function of WD40 domain proteins. Protein Cell 2: 202-214.
- Chen S, Lin F, Hamm HE (2005) RACK1 binds to a signal transfer region of G betagamma and inhibits phospholipase C beta2 activation. J Biol Chem 280: 33445-33452.
- Yaka R, Thornton C, Vagts AJ, Phamluong K, Bonci A, et al. (2002) NMDA receptor function is regulated by the inhibitory scaffolding protein, RACK1. Proc Natl Acad Sci USA 99: 5710-5715.
- Yatime L, Hein KL, Nilsson J, Nissen P (2011) Structure of the RACK1 Dimer from Saccharomyces cerevisiae. J Mol Biol 411: 486-498.
- Rabl J, Leibundgut M, Ataide SF, Haag A, Ban N (2011) Crystal Structure of the Eukaryotic 40S Ribosomal Subunit in Complex with Initiation Factor 1. Science 331: 730-736.
- Adams DR, Ron D, Kiely PA (2011) RACK1, A multifaceted scaffolding protein: Structure and function. Cell Comm Signal 9(22): 823-830.
- Guo J, Chen JG, Liang JS (2007) RACK1, a Versatile Scaffold Protein in Plant? Internl J Plant Dev Biol 1: 95-105.
- Carrillo DR, Chandrasekaran R, Nilsson M, Cornvik T, et al. (2012) Structure of human Rack1 protein at a resolution of 2.45Å. Acta Crystallogr Sect F Struct Biol Cryst Commun 68: 867-872.
- Ullah H, Scappini EL, Moon AF, Williams LV, et al. (2008) Structure of a signal transduction regulator, RACK1, from Arabidopsis thaliana. Protein Sci 17: 1771-80.
- Li D, Roberts R (2001) WD-repeat proteins: structure characteristics, biological function, and their involvement in human diseases. Cell Mol Life Sci 58: 2085-2097.
- McCahill A, Warwicker J, Bolger GB, Houslay MD, Yarwood SJ (2002) The RACK1 scaffold protein: a dynamic cog in cell response mechanisms. Mol Pharm 62: 1261-1273.
- Neer EJ, Schmidt CJ, Nambudripad R, Smith TF (1994) The ancient regulatory-protein family of WD-repeat proteins. Nature 371: 297-300.
- Nilsson J, Sengupta J, Frank J, Nissen P (2004) Regulation of eukaryotic translation by the RACK1 protein: a platform for signaling molecules on the ribosome. EMBO Rep 5: 1137-41.
- Sklan EH, Podoly E, Soreq H (2006) RACK1 has the nerve to act: structure meets function in the nervous system. Prog Neurobiol 78:117-34.
- Nakashima A, Chen L, Thao NP, Fujiwara M, Wong HL, et al. (2008) RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 20: 2265-2279.
- Guillemot F, Billault A, Auffray C (1989) Physical linkage of a guanine nucleotide-binding protein-related gene to the chicken major histocompatibility complex. Proc Natl Acad Sci USA 86: 4594-4598.
- Ishida S, Takahashi Y, Nagata T (1993) Isolation of cDNA of an auxinregulated gene encoding a G protein beta subunit-like protein from tobacco BY-2 cells. Proc Natl Acad Sci USA 90: 11152-11156.
- Ishida S, Takahashi Y, Nagata T (1996) The mode of expression and promoter analysis of the arcA gene, an auxin-regulated gene in tobacco BY-2 cells. Plant Cell Physiol 37: 439-448.
- Iwasaki Y, Komano M, Ishikawa A, Sasaki T, Asahi T (1995) Molecular cloning and characterization of cDNA for a rice protein that contains seven repetitive segments of the Trp-Asp forty-amino-acid repeat (WD-40 repeat). Plant Cell Physiol 36: 505-510.
- McKhann HI, Frugier F, Petrovics G, de la Pena TC, Jurkevitch E, et al. (1997) Cloning of a WD-repeat-containing gene from alfalfa (Medicago sativa): a role in hormone-mediated cell division? Plant Mol Biol 34: 771-80.
- Vahlkamp LPK (1997) AtArcA. Accession No. U77381, the Arabidopsis thaliana homolog of the tobacco ArcA gene (PGR97-145). Plant Physiol 115: 863.
- Schloss JA (1990) A Chlamydomonas gene encodes a G-protein β-subunit like polypeptide. Mol Gen Genet 221: 443-452.
- Wang S, Chen JZ, Zhang Z, Gu S, Ji C, et al. (2003) Cloning, expression and genomic structure of a novel human GNB2L1 gene, which encodes a receptor of activated protein kinase C (RACK). Mol Biol Rep 30: 53-60.
- Nielsen NC, Beilinson V, Bassüner R, Reverdatto S (2001) A Gβ -like protein from soybean. Physiol Plant 111: 75-82.
- Dalrymple MA, Petersen Bjorn S, Friesen JD, Beggs JD (1989) The product of the PRP4 gene of S. cerevisiae shows homology to β subunits of G proteins. Cell 58: 811-812.
- Smith TF, Gaitatzes C, Saxena K, Neer EJ (1999) The WD repeat: a common architecture for diverse functions. Trends Biochem Sci 24: 181-185.
- Sondek J, Bohm A, Lambright DG, Hamm HE, Sigler PB (1996) The 2.1 Å crystal structure of a G protein βγ dimer. Nature 379: 369-374.
- Lambright DG, Sondek J, Bohm A, Skiba NP, Hamm HE, Sigler PB (1996) The 2.0 A crystal structure of a heterotrimeric G protein. Nature 379: 311-319.
- Coyle SM, Gilbert WV, Doudna JA (2009) Direct link between RACK1 function and localization at the ribosome in vivo. Mol Cell Biol 29: 1626-1634.
- Kiely PA, Baillie GS, Lynch MJ, Houslay MD, O’Connor R (2008) Tyrosine 302 in RACK1 is essential for insulin-like growth factor-I-mediated competitive binding of PP2A and beta1 integrin and for tumor cell proliferation and migration. J Biol Chem 283: 22952-22961.
- Kiely PA, O’Gorman D, Luong K, Ron D, O’Connor R (2006) Insulin-like growth factor I controls a mutually exclusive association of RACK1 with protein phosphatase 2A and beta1 integrin to promote cell migration. Mol Cell Biol 26: 4041-4051.
- Stebbins EG, Mochly-Rosen D (2001) Binding specificity for RACK1 resides in the V5 region of beta II protein kinase C. J Biol Chem 276: 29644-29650.
- Mamidipudi V, Zhang J, Lee KC, Cartwright CA (2004) RACK1 regulates G1/S progression by suppressing Src kinase activity. Mol Cell Biol 24: 6788-6798.
- Chen JG, Ullah H, Temple B, Liang J, Guo J, et al. (2006) RACK1 mediates multiple hormone responsiveness and developmental processes in Arabidopsis. J Exp Bot 57: 2697-2708.
- Guo J, Wang J, Xi L, Huang WD, Liang J, Chen JG (2009) RACK1 is a negative regulator of ABA responses in Arabidopsis. J Exp Bot 60: 3819-3833.
- Islas-Flores T, Guillén G, Islas-Flores I, Román-Roque CS, Sánchez F, et al. (2009) Germination behavior, biochemical features and sequence analysis of the RACK1/arcA homolog from Phaseolus vulgaris. Physiol Plant 137: 264-280.
- Li DH, Liu H, Yang YL, Zhen PP, Liang JS (2008) Down-Regulated Expression of RACK1 Gene by RNA Interference Enhances Drought Tolerance in Rice. Chin J Rice Sci 22:447-453.
- Guo J, Chen JG (2008) RACK1 genes regulate plant development with unequal genetic redundancy in Arabidopsis. BMC Plant Biol 8: 108.
- Islas-Flores T, Guillén G, Sánchez F, Villanueva MA (2012) Changes in RACK1 expression induce defects in nodulation and development in Phaseolus vulgaris. Plant Sig Behav 7: 1-3.
- Boyer JS (1982) Plant Productivity and Environment Science 218: 443-448.
- Kundu N, Dozier U, Deslandes L, Somssich IE, Ullah H (2013) Arabidopsis scaffold protein RACK1A interacts with diverse environmental stress and photosynthesis related proteins. Plant Signal Behav 8: e24012.
- Shi S, Chen W, Sun W (2011) Comparative proteomic analysis of the Arabidopsis cbl1 mutant in response to salt stress. Proteomics 11: 4712-4725.
- Kawasaki T, Henmi K, Ono E, Hatakeyama S, Iwano M, et al. (1999) The small GTP-binding protein Rac is a regulator of cell death in plants. Proc Natl Acad Sci USA 96:10922-10926.
- Nakashima A, Chen L, Thao NP, Fujiwara M, Wong HL, et al. (2008) RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 20: 2265-2279.
- Wamaitha MJ, Yamamoto R, Wong HL, Kawasaki T, Kawano Y, et al. (2012) OsRap2.6 transcription factor contributes to rice innate immunity through its interaction with Receptor for Activated Kinase-C 1 (RACK1). Rice 5:35.
- Sengupta J, Nilsson J, Gursky R, Spahn CM, Nissen P, et al. (2004) Identification of the versatile scaffold protein RACK1 on the eukaryotic ribosome by cryo-EM. Nature Struct Mol Biol 11: 957-962.
- Chang IF, Szick-Miranda K, Pan S, Bailey-Serres J (2005) Proteomic characterization of evolutionarily conserved and variable proteins of Arabidopsis cytosolic ribosomes. Plant Physiol 137: 848-862.
- Manuell AL, Yamaguchi K, Haynes PA, Milligan RA, Mayfield SP (2005) Composition and structure of the 80S ribosome from the green alga Chlamydomonas reinhardtii: 80S ribosomes are conserved in plants and animals. J Mol Biol 351: 266-279.
- Giavalisco P, Wilson D, Kreitler T, Lehrach H, Klose J, et al. (2005) High heterogeneity within the ribosomal proteins of the Arabidopsis thaliana 80S ribosome. Plant Mol Biol 57: 577-591.
- Hummel M, Cordewener JHG, de Groot JCM, Smeekens S, America AHP, et al. (2012) Dynamic protein composition of Arabidopsis thaliana cytosolic ribosomes in response to sucrose feeding as revealed by label free MSE proteomics. Proteomics 12: 1024-1038.
- Tan J, Wang J, Chai T, Zhang Y, Feng S, et al. (2013) Functional analyses of TaHMA2, a P1B-type ATPase in wheat. Plant Biotechnol J 11: 420-431.
- Link AJ, Eng J, Schieltz DM, Carmack E, Mize GJ, et al. (1999) Direct analysis of protein complexes using mass spectrometry. Nat Biotech 17: 676-682.
- Shor B, Calaycay J, Rushbrook J, McLeod M (2003) Cpc2/RACK1 is a ribosome-associated protein that promotes efficient translation in Schizosaccharomyces pombe. J Biol Chem 278: 49119-49128.
- Ceci M, Gaviraghi C, Gorrini C, Sala LA, Offenhauser N, et al. (2003) Release of elF6 (p27-BBP) from the 60S subunit allows 80S ribosome assembly. Nature 426: 579-584.
- Jannot G, Bajan S, Giguère NJ, Bouasker S, Banville IH, et al. (2011) The ribosomal protein RACK1 is required for microRNA function in both C.elegans and humans. EMBO reports 12: 581-586.
- Dell EJ, Connor J, Chen S, Stebbins EG, Skiba NP, et al. (2002) The betagamma subunit of heterotrimeric G proteins interacts with RACK1 and two other WD repeat proteins. J Biol Chem 277: 49888-49895.
- Chen S, Dell EJ, Lin F, Sai J, Hamm HE (2004) RACK1 regulates specific functions of Gbetagamma. J Biol Chem 279: 17861-17868.
- Chen S, Spiegelberg BD, Lin F, Dell EJ, Hamm HE (2004) Interaction of Gbetagamma with RACK1 and other WD40 repeat proteins. J Mol Cell Cardiol 37: 399-406.
- Komatsu S, Abbasi F, Kobori E, Fujisawa Y, Kato H, et al. (2005) Proteomic analysis of rice embryo: an approach for investigating Galpha proteinregulated proteins. Proteomics 5: 3932-3941.
- Olejnik K, Bucholc M, Anielska-Mazur A, Lipko A, Kujawa M, et al. (2011) Arabidopsis thaliana Nudix hydrolase AtNUDT7 forms complexes with the regulatory RACK1A protein and Ggamma subunits of the signal transducing heterotrimeric G protein. Acta Biochimica Pol 58: 609-616.