
MicroRNA-124 Suppresses Tumor Cell Proliferation and Invasion by Targeting CD164 Signaling Pathway in Non-Small Cell Lung Cancer
Jing Lin1, Kai Xu1, Jun Wei2, Amy B Heimberger2, Jack A. Roth1 and Lin Ji1*
- 1Department of Thoracic & Cardiovascular Surgery, University of Texas MD Anderson Cancer Center, USA
- 2Department of Neurosurgery, University of Texas MD Anderson Cancer Center, USA
*Address for Correspondence Lin Ji, Department of Thoracic & Cardiovascular Surgery, University of Texas MD Anderson Cancer Center, Houston, TX, USA, Tel: 7135639143; Fax: 7137928905; E-mail: lji@mdanderson.org
Citation: Lin J, Xu K, Wei J, Heimberger AB, Jack AR, et al. MicroRNA-124 Suppresses Tumor Cell Proliferation and Invasion by Targeting CD164 Signaling Pathway in Non-Small Cell Lung Cancer. J Gene Ther 2016; 2(1): 6.
Copyright © 2016 Lin J, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Gene Therapy | ISSN: 2330-2178 | Volume: 2, Issue: 1
Submission:04 January, 2016 | Accepted: 9 February, 2016 | Published: 13 February, 2016
Abstract
MicroRNAs play critical roles in regulating gene expression andvarious cellular processes in human cancer malignant progression. Down-regulated expression of miR-124 gene has been shown to be significantly associated with a poor prognosis in patients with non-small cell lung cancer (NSCLC) but its biological function and regulatory roles in lung cancer tumorigenesis are largely unknown. In this study, we aimed to determine effects of ectopic expression of miR-124 on tumor cell proliferation, invasion, and induction of apoptosis by DOTAP: Cholesterol nanoparticle-mediated gene transfer and identify its endogenous targets under physiological conditions in NSCLC cells. Over-expression of miR-124 significantly suppresses tumor cell proliferation, colony formation, migration, and induction of apoptosis in H322 and A549 cells. Two endogenous miR-124 targeting sites in the 3’ UTR of CD164 mRNA are identified by a stem-loop-array reverse transcription PCR (SLA-RT-PCR) assay in H1299 cells under physiological condition. Ectopic expression of miR-124 induces CD164 mRNA cleavage and down-regulated its gene and protein expression. Our results suggest that miR-124 function as a tumor suppressor miRNA and suppress tumor proliferation and aggression by directly targeting oncogenic CD164 signaling pathway in NSCLC.Introduction
MicroRNAs (miRNAs) are a class of highly conserved small RNA molecules that function as critical regulators of gene expressionthrough binding to the 3’-untranslated region (3’-UTR) of target mRNAs resulting in either mRNA degradation or translation inhibition [1-3]. MicroRNAs are initially transcribed as long primary transcripts (pri-miRNAs) that undergo sequential processing by the RNase III endonucleases Drosha and Dicer to yield the mature 20-23 nucleotide species [4]. Mature miRNAs associate with the RNA induced silencing complex (RISC) and interact with their binding sites with imperfect complementarity in 3’ untranslated regions (UTRs) of target mRNAs. Targeted transcripts subsequently undergo accelerated degradation and reduction of protein production [4,5]. It has been estimated that miRNAs may regulate one third to as many as two thirds of human and mammalian genes and may function as oncogenes or tumor suppressors depending on their targets [6-9]. A growing body of evidence has shown that miRNAs are essential for normal cellular function and development and dysfunction of miRNAs has been linked to many human diseasesand cancer pathogenesis [10].Materials and Methods
To construct and optimize three human precursor miR-124 (premiR-124) gene expressing plasmids, 86- nucleotides of pre-miR124 1(caggcctctctctccgtgttcacagcggaccttgatttaaatgtccatacaattaaggcacgcggtgaatgccaagaatggggctg), 109-nucleotides of pre-miR124-2 (atcaagattagaggctctgctctccgtgttcacagcggaccttgatttaatgtcatacaattaaggcacgcggtgaatgccaagagcggagcctacggctgcacttgaa), and 87-nucleotides of pre-miR124-3 (tgagggcccctctgcgtgttcacagcggaccttgatttaatgtctatacaattaaggcacgcggtgaatgccaagagaggcgcctcc) DNAs were synthesized from Sigma. Seven plasmids were constructed with the same clinically-proven backbone consisting of an expression cassette with a CMV promoter and BGH polyA signaling sequences [34] and a combination of three miR-124 precursor sequences in tandem or individually, and they were named as pre-miR124-123, pre-miR124-1, pre-miR124-2, pre-miR124-3, pre-miR124-12, pre-miR124-23, premiR124-13, as illustrated in Figure 1. All constructs were confirmed by DNA sequencing.
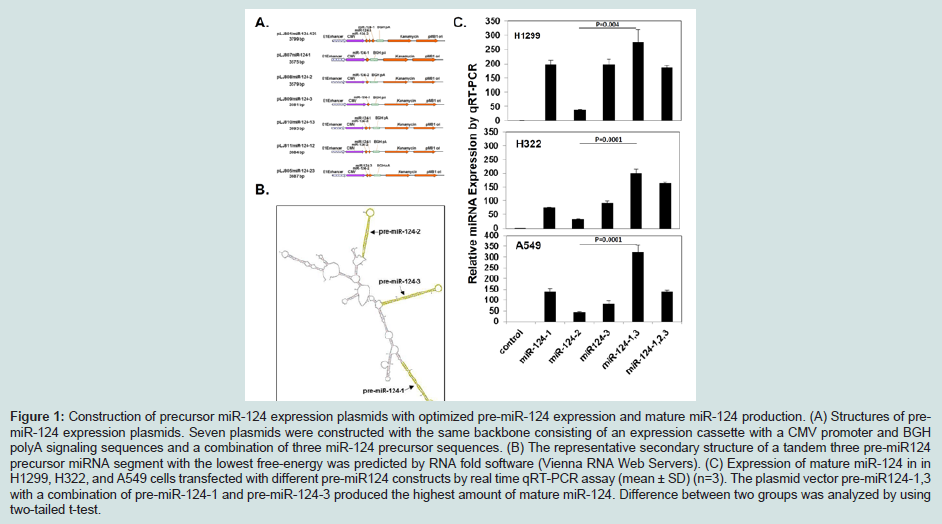
Results
Differential expression of miRNA-124 by different premiR-124 plasmid constructs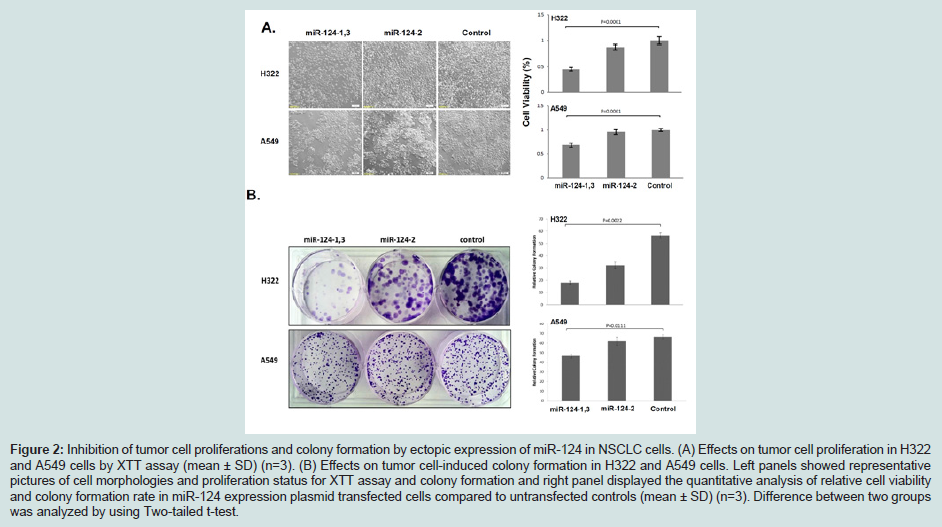
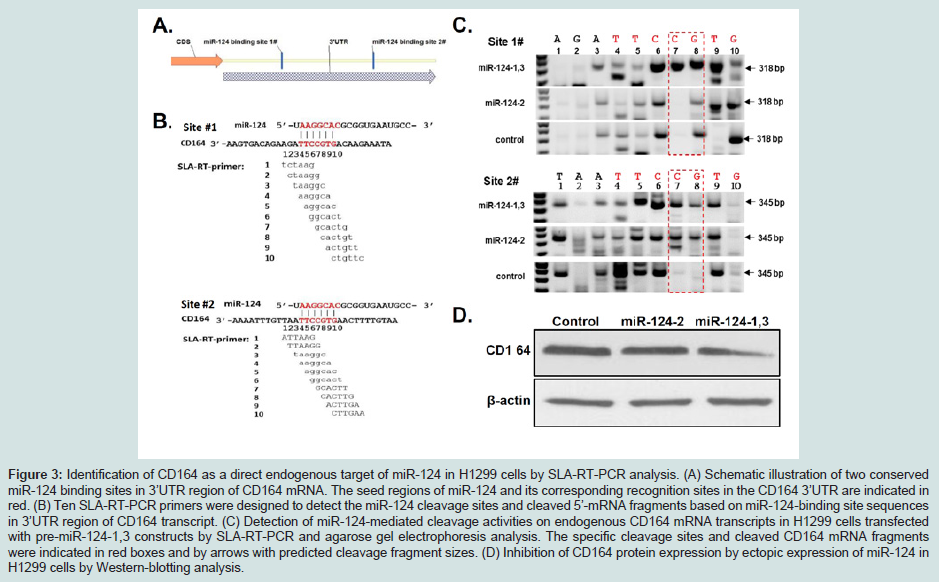
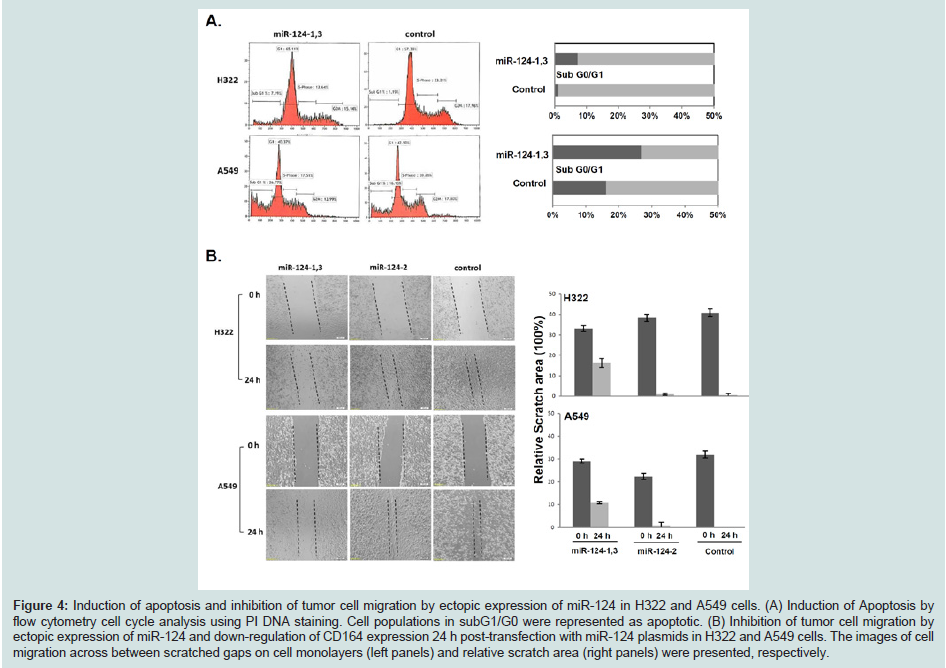
Discussion
MicroRNA-based anticancer therapies are being developed to target multiple cancer-driven effector genes in tumor suppressing and oncogenic signaling pathways involved in cell differentiation, proliferation, malignant progression, and survival [37]. Overexpression of miRNAs by introduction of pre-miRNAs plasmids into cells has various advantages compared to synthetic miR mimics. However, pre-miRNAs expression cassettes have not been fully optimized by current studies. Pre-miRNA-124 is encoded in three distinct chromosomal regions but expresses the identical mature form of mature miRNA-124. Because of different secondary structure and flanking sequences in three pre-miR-124 precursorsthe overexpression of pre-miR-124-2 produced the lowest amount of mature miR-124, while pre-miR-124-1,3 plasmid construct containing both the pre-miR-124-1 and pre-miR-124-3 resulted in the highest amount of mature miR-124 production. Our study showed that individual pre-miRNA constructs produce mature miRNAs in different efficiency, emphasizing the importance to optimize miRNA expression constructs for appropriate therapeutic applications of miRNAs. There are two approaches to re-introduce miRNAs into target cells: transfection of synthetic miRNAs (miRNA mimics) and miRNA-expressing vectors. Transfection of synthetic miRNAs is easy and convenient, however, the lack of in vivo efficient delivery and extremely high cost of miRNA mimics synthesis limits their therapeutic use [37]. In contrast, miRNA-expressing vectors have advantages over synthetic miRNA mimics for its unlimited expression of miRNAs and availability of clinically approved DC-nanoparticlemediated systemic administration, which has been developed in our laboratory and successfully used to delivery therapeutic tumor suppressor genes in preclinical and human clinical trials in NSCLC patients [34].Acknowledgements
This work is supported in part by the National Institutes of Health/National Cancer Institute through a Specialized Program of Research Excellence (SPORE) Grant CA-070907 (J. Minna), and CA176568-02 (Ji, Roth, Wu), a Cancer Prevention and Research Institute of Texas (CPRIT) grant RP130502 (Wu), a MD Anderson’s Cancer Center Support Grant CA-016672-Lung Program, DNA Analysis Facility Shared Resource.
References
- Kato M, Slack FJ (2008) microRNAs: small molecules with big roles - C. elegans to human cancer. Biol Cell 100: 71-81.
- Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, et al. (2005) Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 433: 769-773.
- Calin GA, Croce CM (2006) MicroRNA signatures in human cancers. Nat Rev Cancer 6: 857-866.
- Lee Y, Kim M, Han J, Yeom KH, Lee S, et al. (2004) MicroRNA genes are transcribed by RNA polymerase II. EMBO J 23: 4051-4060.
- Valencia-Sanchez MA, Liu J, Hannon GJ, Parker R (2006) Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev 20: 515-524.
- Lewis BP, Burge CB, Bartel DP (2005) Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 120: 15-20.
- Friedman RC, Farh KK, Burge CB, Bartel DP (2009) Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 19: 92-105.
- Garzon R, Calin GA, Croce CM (2009) MicroRNAs in cancer. Annu Rev Med 60: 167-179.
- Garzon R, Fabbri M, Cimmino A, Calin GA, Croce CM (2006) MicroRNA expression and function in cancer. Trends Mol Med 12: 580-587.
- Kloosterman WP, Plasterk RH (2006) The diverse functions of microRNAs in animal development and disease. Dev Cell 11: 441-450.
- Feng T, Xu D, Tu C, Li W, Ning Y, et al. (2015) MiR-124 inhibits cell proliferation in breast cancer through downregulation of CDK4. Tumour Biol 36: 5987-5997.
- Li X, Yu Z, Li Y, Liu S, Gao C, et al. (2015) The tumor suppressor miR-124 inhibits cell proliferation by targeting STAT3 and functions as a prognostic marker for postoperative NSCLC patients. Int J Oncol 46: 798-808.
- Jeong D, Kim J, Nam J, Sun H, Lee YH, et al. (2015) MicroRNA-124 links p53 to the NF-kB pathway in B-cell lymphomas. Leukemia 29: 1868-1874.
- Wei J, Wang F, Kong LY, Xu S, Doucette T, et al. (2013) miR-124 inhibits STAT3 signaling to enhance T cell-mediated immune clearance of glioma. Cancer Res 73: 3913-3926.
- An L, Liu Y, Wu A, Guan Y (2013) microRNA-124 inhibits migration and invasion by down-regulating ROCK1 in glioma. PLoS One 8: e69478.
- Cai JJ, Qi ZX, Chen LC, Yao Y, Gong Y, et al. (2016) miR-124 suppresses the migration and invasion of glioma cells in vitro via Capn4. Oncol Rep 35: 284-290.
- Berghmans T, Ameye L, Willems L, Paesmans M, Mascaux C, et al. (2013) Identification of microRNA-based signatures for response and survival for non-small cell lung cancer treated with cisplatin-vinorelbine A ELCWP prospective study. Lung Cancer 82: 340-345.
- Guo W, Zhang Y, Zhang Y, Shi Y, Xi J, et al. (2015) Decreased expression of miR-204 in plasma is associated with a poor prognosis in patients with non-small cell lung cancer. Int J Mol Med 36: 1720-1726.
- 19. Sun Y, Ai X, Shen S, Lu S (2015) NF-ĸB-mediated miR-124 suppresses metastasis of non-small-cell lung cancer by targeting MYO10. Oncotarget 6: 8244-8254.
- 20. Wu S, Shen W, Pan Y, Zhu M, Xie K, et al. (2015) Genetic variations in key microRNAs are associated with the survival of nonsmall cell lung cancer. Medicine (Baltimore) 94: e2084.
- Zhang Y, Li H, Han J, Zhang Y (2015) Down-regulation of microRNA-124 is correlated with tumor metastasis and poor prognosis in patients with lung cancer. Int J Clin Exp Pathol 8: 1967-1972.
- Li Y, Zhang Z, Liu X, Huang T, He W, et al. (2014) miR-124 functions as a tumor suppressor in the endometrial carcinoma cell line HEC-1B partly by suppressing STAT3. Mol Cell Biochem 388: 219-231.
- Xu X, Li S, Lin Y, Chen H, Hu Z, et al. (2013) MicroRNA-124-3p inhibits cell migration and invasion in bladder cancer cells by targeting ROCK1. J Transl Med 11: 276.
- Hu CB, Li QL, Hu JF, Zhang Q, Xie JP, et al. (2014) miR-124 inhibits growth and invasion of gastric cancer by targeting ROCK1. Asian Pac J Cancer Prev 15: 6543-6546.
- Shi XB, Ma AH, Xue L, Li M, Nguyen HG, et al. (2015) mir-124 and androgen receptor signaling inhibitors repress prostate cancer growth by downregulating androgen receptor splice variants, EZH2, and Src. Cancer Res 75: 5309-5317.
- Jiang L, Lin T, Xu C, Hu S, Pan Y, et al. (2016) miR-124 interacts with the Notch1 signalling pathway and has therapeutic potential against gastric cancer. J Cell Mol Med 20: 313-322.
- Zhang T, Wang J, Zhai X, Li H, Li C, et al. (2014) MiR-124 retards bladder cancer growth by directly targeting CDK4. Acta Biochim Biophys Sin (Shanghai) 46: 1072-1079.
- Peng XH, Huang HR, Lu J, Liu X, Zhao FP, et al. (2014) MiR-124 suppresses tumor growth and metastasis by targeting Foxq1 in nasopharyngeal carcinoma. Mol Cancer 13: 186.
- Zheng F, Liao YJ, Cai MY, Liu YH, Liu TH, et al. (2012) The putative tumour suppressor microRNA-124 modulates hepatocellular carcinoma cell aggressiveness by repressing ROCK2 and EZH2. Gut 61: 278-289.
- Xia J, Wu Z, Yu C, He W, Zheng H, et al. (2012) miR-124 inhibits cell proliferation in gastric cancer through down-regulation of SPHK1. J Pathol 227: 470-480.
- Lang Q, Ling C (2012) MiR-124 suppresses cell proliferation in hepatocellular carcinoma by targeting PIK3CA. Biochem Biophys Res Commun 426: 247-252.
- Hunt S, Jones AV, Hinsley EE, Whawell SA, Lambert DW (2011) MicroRNA-124 suppresses oral squamous cell carcinoma motility by targeting ITGB1. FEBS Lett 585: 187-192.
- Zhang H, Wang Q, Zhao Q, Di W (2013) MiR-124 inhibits the migration and invasion of ovarian cancer cells by targeting SphK1. J Ovarian Res 6: 84.
- Lu C, Stewart DJ, Lee JJ, Ji L, Ramesh R, et al. (2012) Phase I clinical trial of systemically administered TUSC2 (FUS1)-nanoparticles mediating functional gene transfer in humans. PLoS One 7: e34833.
- Lin J, Xu K, Gitanjali J, Roth JA, Ji L (2011) Regulation of tumor suppressor gene FUS1 expression by the untranslated regions of mRNA in human lung cancer cells. Biochem Biophys Res Commun 410: 235-241.
- Suzhi Z, Liang T, Yuexia P, Lucy L, Xiaoting H, et al. (2015) Gap junctions enhance the antiproliferative effect of microRNA-124-3p in glioblastoma cells. J Cell Physiol 230: 2476-2488.
- Garzon R, Marcucci G, Croce CM (2010) Targeting microRNAs in cancer: rationale, strategies and challenges. Nat Rev Drug Discov 9: 775-789.
- Tang J, Zhang L, She X, Zhou G, Yu F, et al. (2012) Inhibiting CD164 expression in colon cancer Cell line HCT116 leads to reduced cancer cell proliferation, mobility, and metastasis in vitro and in vivo. Cancer Invest 30: 380-389.