
Journal of Vaccine & Immunotechnology
Download PDF
Research Article
*Address for Correspondence: Toshihiro Maekawa, Institute for Health Care Science, Suntory Wellness Limited, 1-1-1 Wakayama-dai, Shimamoto-cho, Mishimagun, Osaka 618-8503, Japan, Tel: +81-75-962-6207; Fax: +81-75-962-1690; E-mail: Toshihiro_Maekawa@suntory.co.jp
Citation: Maekawa T, Kimoto T, Mizuno D, Furukawa Y, Ida M, et al. Oral Administration of Lactobacillus pentosus Strain S-PT84 Enhances Anti-Influenza Virus-Specific IgG Production in Plasma after Limited Doses of Influenza Virus Vaccination in Mice. J Vaccine Immunotechnology. 2015;2(1): 5.
Copyright © 2014 Maekawa T, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Vaccine & Immunotechnology | Volume: 2, Issue: 1
Submission: 27 November 2014 | Accepted: 23 December 2014 | Published: 29 December 2014
Reviewed & Approved by: Dr. Sherven Sharma, Professor of Medicine, David Geffen School of Medicine, University of California, USA.
Keywords: Influenza vaccine; Lactobacillus pentosus strain S-PT84; Adjuvant effect; IgG; IgA
Oral Administration of Lactobacillus pentosus Strain S-PT84 Enhances Anti-Influenza Virus-Specific IgG Production in Plasma after Limited Doses of Influenza Virus Vaccination in Mice
Toshihiro Maekawa1*, Takashi Kimoto2, Dai Mizuno2, Yuichi Furukawa1, Masayuki Ida1, Etsuhisa Takahashi2, Takayuki Izumo1, Yoshiko Ono1, Hiroshi Shibata1 and Hiroshi Kido2
- 1Institute for Health Care Science, Suntory Wellness Limited, Mishima-gun, Osaka, Japan
- 22Division of Enzyme Chemistry, Institute for Enzyme Research, The University of Tokushima, Tokushima, Japan
*Address for Correspondence: Toshihiro Maekawa, Institute for Health Care Science, Suntory Wellness Limited, 1-1-1 Wakayama-dai, Shimamoto-cho, Mishimagun, Osaka 618-8503, Japan, Tel: +81-75-962-6207; Fax: +81-75-962-1690; E-mail: Toshihiro_Maekawa@suntory.co.jp
Citation: Maekawa T, Kimoto T, Mizuno D, Furukawa Y, Ida M, et al. Oral Administration of Lactobacillus pentosus Strain S-PT84 Enhances Anti-Influenza Virus-Specific IgG Production in Plasma after Limited Doses of Influenza Virus Vaccination in Mice. J Vaccine Immunotechnology. 2015;2(1): 5.
Copyright © 2014 Maekawa T, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Vaccine & Immunotechnology | Volume: 2, Issue: 1
Submission: 27 November 2014 | Accepted: 23 December 2014 | Published: 29 December 2014
Reviewed & Approved by: Dr. Sherven Sharma, Professor of Medicine, David Geffen School of Medicine, University of California, USA.
Abstract
Background: It has been reported that various Lactobacillus species enhance antigen-specific antibody production after viral infection and/or vaccination in animals and humans. In this study, the effect of oral administration of Lactobacillus pentosus strain S-PT84 on subcutaneous administration of limited doses of 2.5 ng of influenza A virus (IAV) (H1N1) split hemagglutinin (HA) vaccine was studied in mice.Methods: Seven-week-old BALB/c female mice were fed the AIN-93M diet with or without 0.186% S-PT84 for 6 weeks ad libitum and then given subcutaneous injection of IAV/California/7/2009 (H1N1) HA vaccine at doses of 2.5–15.0 ng/mice and boosted on day 28. Two weeks after the last vaccination, the mice were sacrificed under anesthesia, and the amount of anti-HA-specific IgG in plasma was measured by ELISA.
Results: The levels of anti-HA-specific IgG in plasma were significantly higher in the S-PT84 group than in the control group without S-PT84 treatment. Anti-HA-specific IgA levels in nasal washes were under the detection level in both groups.
Conclusion: Oral administration of S-PT84 enhanced the production of HA-specific IgG antibody in plasma after subcutaneous vaccination, even at the limited dose of 2.5 ng of IAV (H1N1) HA in mice, suggesting that S-PT84 has a potent adjuvant activity against IAV HA.
Keywords: Influenza vaccine; Lactobacillus pentosus strain S-PT84; Adjuvant effect; IgG; IgA
Introduction
Influenza is an acute viral infection that results in high morbidity and significant mortality, particularly in infants and the elderly populations [1]. They sometimes have a high risk of fatal influenza pneumonia and thus vaccination is recommended for prevention. The World Health Organization has recommended use of oseltamivir (Tamiflu®, Chugai Pharmaceutical Co., Ltd, Tokyo, Japan) and zanamivir (Relenza®, Glaxo Smith Kline K.K., Tokyo, Japan) for the treatment of influenza. These antiviral neuraminidase inhibitors are useful options for seasonal influenza infections in the world [2]. However, Shinahara et al. recently reported that administration of these inhibitors decreases anti-IAV HA-specific IgG and IgA production in plasma and nasopharyngeal secretions, probably because of inhibition of viral replication and limited production of viral antigens [3]. The limited viral antigens do not induce sufficient acquired immunity in patients treated with anti-viral neuraminidase inhibitors, resulting in a high frequency of influenza virus re-infection in the subsequent year [3].Lactic acid bacteria (LAB) are widely used as a health food ingredient and could modulate mucosal and systemic immune responses [1,5]. Oral administration of live Lactobacillus casei Shirota significantly increases the survival rate of neonatal and infant mice infected with IAV by stimulating IL-12 production and NK activity in the lung [6]. Oral administration of heat-killed Lactobacillus plantrum L-137 [7] and Lactobacillus pentosus strain b240 [8] enhances type 1 interferon production and increases the production of anti-IAV IgG in plasma and IgA in bronchoalveolar lavage fluid, respectively, and prolongs the survival period of mice infected with IAV. These studies showed that oral administration of live and nonviable Lactobacillus species enhances the protective effects against airway infection of IAV
It has recently been reported that oral administration of probiotic LAB enhances vaccination-induced antibody production. Lactobacillus acidophilus enhances rotavirus-specific antibody after vaccination of rotavirus in the neonatal gnotobiote pig [9]. Davidson et al. reported that volunteers receiving Lactobacillus GG show higher protective titers after administration of live attenuated IAV/ H3N2 vaccine compared to a placebo group after vaccination [10]. Moreover, Boge et al. reported that daily consumption of a probiotic drink increases relevant specific antibody responses to influenza vaccination in healthy volunteers over 70 years of age [11]. Therefore oral administration of Lactobacillus species might be effective in preventing influenza virus infection by stimulating antibody production.
We previously reported that Lactobacillus pentosus strain S-PT84 enhances splenic natural killer (NK) activity and exhibits anti-allergic effects by modulating T-helper1/T-helper2 (Th1/Th2) balance [12]. Moreover, it has been reported that oral administration of S-PT84 enhances interferon-α production from plasmacytoid dendritic cells against an insufficient dose of hemagglutinating virus of Japan (HVJ) infection [13], and that intranasal administration of S-PT84 protects against influenza infection [14]. However, the effects of S-PT84 ingestion on limited doses of IAV HA vaccination and HAspecific antibody induction have not yet been examined. In this study, the effect of oral administration of S-PT84 on the induction of IAV HA-specific antibody in mice immunized by IAV HA vaccine in the nanogram range was investigated.

Effect of Lactobacillus pentosus S-PT84 on the production of HA-specific IgG in plasma
Materials and Methods
AnimalsBALB/c female mice, 7 weeks old, were obtained from Japan SLC, Inc. (Shizuoka, Japan). Mice were housed and fed a commercial diet (CE-2) and tap water ad libitum for 1 week at 25±1 oC and 60% ± 5% humidity under a 12 h light-dark cycle before experimentation. Experiments were performed according to the Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, 1996).
Bacterial strains and bacteria preparation
S-PT84 was isolated from Kyoto pickles “SHIBAZUKE” [12] and was cultivated in a medium containing glucose and yeast extract (AromildTM, SK yeast extract Hi-K) at 37 oC for 24 hours. Cultured bacteria were collected by centrifugation at 9,190 x g for 5 min, washed twice with sterile saline and once with distilled water, and heat-killed at 95 oC for 5 min. Heat-killed S-PT84 was lyophilized for use in the experiments.
Vaccine
The influenza antigen used in the present studies, IAV/ California/7/2009(H1N1) virus processed for HA vaccine (splitproduct, 0.636 μg protein/0.341 μg HA/mL), was purchased from DENKA SEIKEN Co., Ltd. (Tokyo, Japan).
Procedure and sample collection
BALB/c mice were fed on a commercial diet (AIN-93M, Oriental Yeast Co. Ltd., Tokyo, Japan) with or without 0.186% S-PT84 and tap water ad libitum for 6 weeks. Mice were given subcutaneous vaccinations (2.5–15.0 ng HA/100 μL) and boosted on day 28. Two weeks after the last vaccination, the mice were sacrificed under anesthesia. Blood samples were collected from the vein. Blood was centrifuged (2,500 g, 10 min) at 4 oC, and the supernatant was collected.
Enzyme-linked immunosorbent assay (ELISA)
Anti-influenza HA-specific antibodies in plasma were measured by ELISA [15]. Briefly, 96-well plates (Nunc, Naperville, IL) were coated with HA vaccine and bovine serum albumin (BSA, 0.1 μg/well each) in PBS overnight at 4 oC. After the pre-coating, it was blocked with 1% BSA in 50 mM Tris–HCl (pH 8.0) containing 0.14 M NaCl and 0.05% Tween 20 (TTS) for 1 hour at room temperature. The plasma diluted with TTS containing 1% BSA was added to each well and incubated for 3 hours at room temperature. The plate was washed six times with TTS containing 1% BSA and incubated with goat anti-mouse IgG antibodies conjugated with horseradish peroxidase (Bethyl Laboratories Inc., Montgomery, TX, USA) for 2 hours at room temperature. Color was developed by the addition of TMB substrate (Bethyl Laboratories), according to the instructions provided by the manufacturer. The chromogen produced was measured at 450 nm using a SPECTRA max PLUS384 AutoReader (Molecular Devices, Tokyo, Japan). Antibody titers were defined as the reciprocal of the highest dilution of sample for which the optical density (OD) was at least twice the OD of the negative control samples before vaccination.
Statistical analysis
The significance of differences in values between the control group and S-PT84 group was determined using Student’s t-test. P-values less than 0.05 were considered significant.
Results
Doses of vaccination of IAV/California/7/2009(H1N1)The induced levels of HA-specific IgG antibody in plasma were analyzed to detect the optimal dose of IAV/California/7/2009(H1N1) to evaluate the effect of Lactobacillus pentosus S-PT84 as an adjuvant. HA-specific IgG antibody in plasma was increased by IAV HA vaccination in a dose-dependent manner in the range between 2.5 and 15.0 ng (Figure 1). HA at 2.5 ng, even at the lowest dosage, could induce production of HA-specific IgG in plasma, and that dose may be appropriate for examining the effect of LAB on enhancing HAspecific IgG production in plasma. Therefore, the minimal dose of 2.5 ng HA was used to investigate the adjuvant effect of LAB.
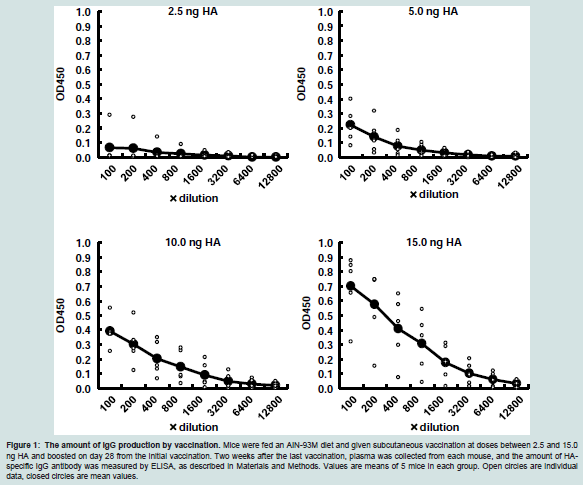
Figure 1: The amount of IgG production by vaccination. Mice were fed an AIN-93M diet and given subcutaneous vaccination at doses between 2.5 and 15.0 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HAspecific IgG antibody was measured by ELISA, as described in Materials and Methods. Values are means of 5 mice in each group. Open circles are individual data, closed circles are mean values.
Effect of Lactobacillus pentosus S-PT84 on the production of HA-specific IgG in plasma
Mice were fed the AIN-93M diet with or without 0.186% S-PT84 for 6 weeks. In the control group treated with two subcutaneous injections of IAV HA at 2.5 ng alone, a small amount of antigenspecific IgG antibody production was detected in plasma. The S-PT84 group of mice treated with continuous feeding of the AIN-93M diet with 0.186% S-PT84 during the experimental period also received two subcutaneous injections of IAV HA at 2.5 ng. HA-specific IgG antibody levels in plasma were higher in the S-PT84 group than in the control group (Figure 2a). On the other hand, HA-specific IgA in nasal washes was under detection levels in both groups (data not shown). The induced HA-specific IgG titers in plasma were significantly higher in the S-PT84 group than in the control group (p=0.007) (Figure 2b).
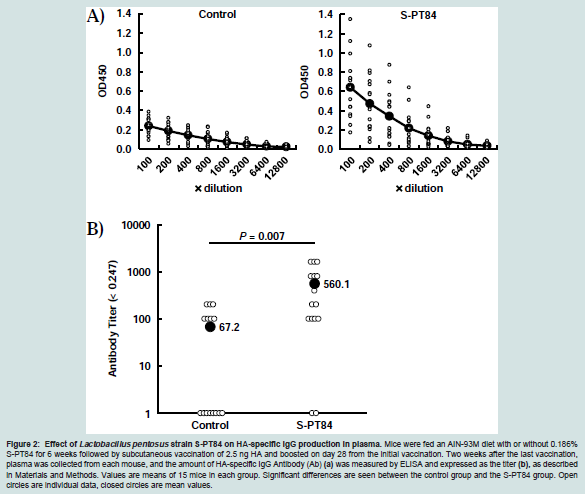
Figure 2: Effect of Lactobacillus pentosus strain S-PT84 on HA-specific IgG production in plasma. Mice were fed an AIN-93M diet with or without 0.186% S-PT84 for 6 weeks followed by subcutaneous vaccination of 2.5 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HA-specific IgG Antibody (Ab) (a) was measured by ELISA and expressed as the titer (b), as describedin Materials and Methods. Values are means of 15 mice in each group. Significant differences are seen between the control group and the S-PT84 group. Open circles are individual data, closed circles are mean values.
Discussion
In this study, oral administration of S-PT84 enhanced HAspecific IgG induction in plasma, but not HA-specific IgA induction in nasal wash, after subcutaneous administration of limited doses of IAV (H1N1) HA in mice. The findings suggest that S-PT84 has an adjuvant effect against IAV HA on plasma IgG induction, even at minimal doses. Nonaka et al. reported that S-PT84 induces cytokine production from peritoneal macrophages in vitro and enhances the activity of NK cells in vivo [12]. Koizumi et al. also reported that S-PT84 stimulates IFN-γ and IL-12 production through Tolllike receptor (TLR)-2 and TLR-4 on dendritic cells [16]. Moreover, Izumo et al. reported that Salmonella-specific antibody is enhanced by S-PT84 ingestion in Salmonella typhimurium-infected mice [17]. Accordingly, we thought that antigen-presenting cells were activated by ingestion of S-PT84 in this experiment, and production of HA-specific IgG in plasma may be enhanced, as in a previous report [17]. It is well known that influenza vaccines administered intramuscularly or subcutaneously induce a predominantly IgGmediated protection in the systemic immune compartment, but this systemic immunization offers inadequate induction of IgA in airway mucosa [18]. Therefore, oral administration of S-PT84 might affect HA antigen-induced dendritic cell-mediated systemic immunity and induce HA-specific IgG in plasma, but not affect mucosal immunity and HA-specific IgA production.The present data suggest that S-PT84 enhances antibody production as an adjuvant with limited doses of influenza viral antigen. Hui-Tsu et al. reported that the induction of HA-specific antibody in plasma was enhanced by IAV HA (H1N1) vaccination at 10 ng dose in mice, and the survival rate was also significantly improved by this vaccination [19]. The optimal density (OD) level of HA-specific IgG was similar compared with our present study. Moreover, the strain of IAV/California/7/2009 (H1N1) was same in both experiments. Therefore, we speculate that HA-specific antibody production level by the killed bacterial adjuvant S-PT84 might be sufficient for the viral protection and then an ample titer was obtained for influenza virus exclusion. It suggests that S-PT84 may decrease the risk of influenza virus re-infection treated with anti-viral neuraminidase inhibitors.
As a next step, it should be to investigate whether oral administration of S-PT84 together with the vaccination on limited dose of HA antigen can protect IAV infection, and/or whether oral administration of S-PT84 can induce sufficiently HA-specific IgG antibody treated with antiviral neuraminidase inhibitors after IAVinfection.
Japan is becoming a super-aging society more rapidly than other countries. Elderly persons have a high risk of influenza-associated pneumonia, which is sometimes fatal. Thus, management of influenza outbreaks is important for them. Indeed, seroconversion and seroprotection are lower in elderly individuals than in adults after influenza vaccination, and their immune functions are weaker than in adults [20]. It is important that preventive action be taken to enhance immune function or provide vaccination for the elderly. In this regard, S-PT84 may have a potential to improve their immune response.
References
- Gillim-Ross L, Subbarao K (2006) Emerging respiratory viruses: Challenges and vaccine strategies. Clin Microbiol Rev 19: 614-636.
- Moscona A (2005) Neuraminidase inhibitors for Influenza. N Engl J Med 353: 1363-1373.
- Shinahara W, Takahashi E, Sawabuchi T, Arai M, Hirotsu N, et al. (2013) Immunomodulator clarithromycin enhances mucosal and systemic immune responses and reduces re-infection rate in pediatric patients with Influenza treated with antiviral neuraminidase inhibitors: A retrospective analysis. PLoS One 8: e70060.
- Blum S, Haller D, Pfeifer A, Schiffrin EJ (2002) Probiotics and immune response. Clin Rev Allergy Immunol 22: 287-309.
- Perdigón G, Fuller R, Raya R (2001) Lactic acid bacteria and their effect on the immune system. Curr Issues Intest Microbiol 2: 27-42.
- Yasui H, Kiyoshima J, Hori T (2004) Reduction of Influenza virus titer and protection against influenza virus infection in infant mice fed Lactobacillus casei Shirota. Clin Diagn Lab Immunol 11: 675-679.
- Maeda N, Nakamura R, Hirose Y, Murosaki S, Yamamoto Y, et al. (2009) Oral administration of heat-killed Lactobacillus plantarum L-137 enhances protection against influenza virus infection by stimulation of type I interferon production in mice. Int Immunopharmacol 9: 1122–1125.
- Kobayashi N, Saito T, Uematsu T, Kishi K, Toba M, et al. (2011) Oral administration of heat-killed Lactobacillus pentosus strain b240 augments protection against influenza virus infection in mice. Int Immunopharmacol 11: 199–203.
- Zhanga W, Azevedoa MSP, Wenb K, Gonzaleza A, Saifa LJ, et al. (2008) Probiotic Lactobacillus acidophilus enhances the immunogenicity of an oral rotavirus vaccine in gnotobiotic pigs. Vaccine 26: 3655–3661.
- Davidson LE, Fiorino AM, Snydman DR, Hibberd PL (2011) Lactobacillus GG as an immune adjuvant for live attenuated Influenza vaccine in healthy adults: a randomized double blind placebo controlled trial. Eur J Clin Nutr 65: 501–507.
- Boge T, Rémigy M, Vaudaine S, Tanguyc J, Bourdet-Sicardc R, et al. (2009) A probiotic fermented dairy drink improves antibody response to influenza vaccination in the elderly in two randomised controlled trials. Vaccine 27: 5677–5684.
- Yuji N, Takayuki I, Fumi I, Toshihiro M, Hiroshi S, et al. (2008) Antiallergic effects of Lactobacillus pentosus strain S-PT84 mediated by modulation of Th1/Th2 immunobalance and induction of IL-10 production. Int Arch Allergy Immunol 145: 249–257.
- Izumo T, Maekawa T, Ida M, Kishi A, Akatani K, et al. (2011) Effect of Lactobacillus pentosus S-PT84 ingestion on IFN-α production from plasmacytoid dendritic cells by virus stimulation. Biosci Biotechnol Biochem 75: 370-372.
- Izumo T, Maekawa T, Ida M, Noguchi A, Kitagawa Y, et al. (2010) Effect of intranasal administration of Lactobacillus pentosus S-PT84 on influenza virus infection in mice. Int Immunopharmacol 10: 1101–1106.
- Nishino M, Mizuno D, Kimoto T, Shinahara W, Fukuta A, et al. (2009) Influenza vaccine with Surfacten, a modified pulmonary surfactant, induces systemic and mucosal immune responses without side effects in minipigs. Vaccine 27: 5620-5627.
- Koizumi S, Wakita D, Sato T, Mitamura R, Izumo T, et al. (2008) Essential role of Toll-like receptors for dendritic cell and NK1.1(+) cell-dependent activation of type 1 immunity by Lactobacillus pentosus strain S-PT84. Immunol Lett 120: 14–19.
- Izumo T, Izumi f, Nakagawa I, Kitagawa Y, Sibata H, et al. (2011) Influence of Lactobacillus pentosus S-PT84 ingestion on the mucosal immunity of healthy and Salmonella typhimurium-infected mice. Biosci Microflora 30: 27–35.
- Kimoto T, Mizuno D, Takei T, Kunimi T, Ono S, et al. (2013) Intranasal influenza vaccination using a new synthetic mucosal adjuvant SF-10: induction of potent local and systemic immunity with balanced Th1 and Th2 responses. Influenza Other Respir Viruses 7: 1218-1226.
- Lin HT, Chuang CC, Wu HL, Chu DM, Wang YC (2013) Characterization of cross protection of Swine-Origin Influenza Virus (S-OIV) H1N1 and reassortant H5N1 influenza vaccine in BALB/c mice given a single-dose vaccination. J Biomed Sci 20: 19.
- Goodwin K, Viboud C, Simonsen L (2006) Antibody response to influenza vaccination in the elderly: A quantitative review. Vaccine 24: 1159–1169.